Презентация “Виды папоротников” – дошкольное образование, презентации
библиотека
материалов
Содержание слайдов
Номер слайда 1
Виды папоротников
Номер слайда 2
Папоротники — одни из немногих древнейших растений, которые сохранили значительное видовое разнообразие, сопоставимое с тем, что было много веков назад. В то время как другие растения исчезали с лица Земли, разные виды папоротников, наоборот, развивались, образуя все новые и новые формы.
В то время как другие растения исчезали с лица Земли, разные виды папоротников, наоборот, развивались, образуя все новые и новые формы.
Номер слайда 3
Адиантум стоповидный. Родиной адиантума стоповидного является Дальний Восток (папоротник растет в лесах Приморья и Приамурья), и восточная часть Северной Америки.
Номер слайда 4
Адиантум Венерин волос. Наиболее распространенным является Венерин волос – папоротник с красивыми ажурными перисто-рассеченными листьями, расположенными на тонких, напоминающих проволоку черешках. Такое название растение заслужило из-за тонких темных стеблей, на которых располагаются маленькие листики-вайи.
Наиболее распространенным является Венерин волос – папоротник с красивыми ажурными перисто-рассеченными листьями, расположенными на тонких, напоминающих проволоку черешках. Такое название растение заслужило из-за тонких темных стеблей, на которых располагаются маленькие листики-вайи.
папоротник страусник. Страусник обыкновенный- впечатляющий по красоте многолетний папоротник из семейства Оноклеевые. В природе произрастает на влажных тенистых лугах и лесах умеренного пояса Северного полушария. В благоприятных условиях вырастает до 150 см в высоту и 100 см в ширину. В народе его часто называют «страусовое перо» или «разнолистник».
В народе его часто называют «страусовое перо» или «разнолистник».
Папоротник пузырник. Пузырник (Cystopteris) представляет собой низкорослый скальный папоротник с ползучими стеблями и узкими удлиненными листьями длиной от 30 до 70 см, в зависимости от вида. Каждый лист делится на множество пар листочков, каждый из которых разделяется на дольчатые сегменты.
Папоротник многорядник. Род многорядника насчитывает около 260 вечнозеленых или полувечнозеленых папоротников из семейства Щитовниковые. Наибольшее видовое разнообразие приходится на Юго-Восточную Азию. Представителей рода можно встретить в Мексике, Бразилии, Африке, Северной Америке и Европе.
Род многорядника насчитывает около 260 вечнозеленых или полувечнозеленых папоротников из семейства Щитовниковые. Наибольшее видовое разнообразие приходится на Юго-Восточную Азию. Представителей рода можно встретить в Мексике, Бразилии, Африке, Северной Америке и Европе.
Номер слайда 8
Гроздовик папоротник Существует всего 8 видов этих реликтовых папоротников, в ландшафтном дизайне используется только два. Один из них, гроздовник виргинский, – достаточно крупное, до 45 см высотой, опушённое растение.
Номер слайда 9
Криптограмма папоротник состав семейства входит около десяти представителей данного вида. Криптограмма имеет изящные хохлатые ярко-зеленые листья, напоминающие петрушку. Распространение: западная и северная часть Северной Америки, север Китая, Монголии, Сибири.
Криптограмма имеет изящные хохлатые ярко-зеленые листья, напоминающие петрушку. Распространение: западная и северная часть Северной Америки, север Китая, Монголии, Сибири.
Номер слайда 10
Папоротник марсилея. Растения семейства марсилея относятся к водным папоротникам. Они растут в мелких и временно пересыхающих водоёмах, культивируются как аквариумная культура. В Германии эти папоротники называют водным клевером, поскольку их округлые четырёхлопастные вайи напоминают листья клевера
Номер слайда 11Орляк обыкновенный. Орляк обыкновенный — растет на всех континентах Земли, образуя заросли в тундре и сухих борах Европы, среди кустарников Австралии и на пустошах Южной Америки, нет его лишь в степях и пустынях.
Орляк обыкновенный — растет на всех континентах Земли, образуя заросли в тундре и сухих борах Европы, среди кустарников Австралии и на пустошах Южной Америки, нет его лишь в степях и пустынях.
Осмунда, чистоуст. Самые крупные древнейшие папоротники умеренной зоны Земли. Некогда росшие на всех материках, они сохранились лишь в лесах Кавказа, Восточной Азии и Северной Америке
Номер слайда 13
Телиптерис болотный.
Номер слайда 14
Фегоптерис. Фегоптерис связывающий (Ph. connectilis = Thelypteris phegopteris) — чаще других папоротников встречается в лесах средней полосы России. Невысокий (до 40 см) папоротник. Благодаря наличию длинного ветвящегося корневища, он образует плотный, быстро разрастающийся напочвенный покров из светло-зеленых дельтовидных листьев на черешках.
Комнатные папоротники
Мир папоротников разнообразен, известно около 300 родов и больше 10000 видов этих растений. Они сильно различаются по размерам, формам, особенностям строения и размножения.
У папоротников нет настоящих листьев. То, что у папоротника напоминает лист – это вайя, целая система ветвей, расположенных в одной плоскости.
Папоротники распространены по всему земному шару, но самое большое разнообразие видов – во влажных тропических лесах. Они растут как эпифиты на деревьях, в расселинах скал, на почве, болотах, возле рек и озёр.
Ботаническая классификация папоротников сложная, иногда запутанная, в литературе один и тот же вид может быть отнесен к разным семействам.
В комнатной культуре обычно выращивают тропические папоротники. Правильный уход в домашних условиях требует обеспечения высокой влажности и тепла для этих растений.
Причем температура не самое главное – есть много видов папоротников, которые не требуют тепла, достаточно зимой +15-20 градусов, а горные виды прекрасно перезимуют и при +10 градусах.
Важно то, что необходимо постоянно поддерживать довольно высокую влажность воздуха, что довольно сложно обеспечить в обычных условиях квартиры в отапливаемый период. Даже ежедневного опрыскивания растений теплой водой не достаточно.
Но проблему решает размещение папоротников рядом с аквариумом или специальным увлажнителем воздуха. Неплохо растут папоротники в зеленом уголке, где собрано в большую композицию много комнатных растений. Их листовые пластинки испаряют влагу, создавая нужный микроклимат в зеленой зоне. Это также хорошее растение для ванных комнат с естественным освещением.
В домашних условиях выращивают наиболее устойчивые виды – Адиантум (Венерины волосы), Асплениум, Нефролепис, Циртомиум, Птерис, Пеллея. Реже встречаются Платицериум (Оленьи рога), Даваллия.
Разные виды папоротников очень отличаются по внешнему виду, но требования к условиям их содержания похожи:
- Любят хорошее рассеянное освещение, но нуждаются в защите от прямых солнечных лучей. Для содержания папоротников лучше всего подходят западные или восточные окна, где солнце светит несколько часов утром или вечером.
- Не переносят холодных сквозняков, но нуждаются в регулярном проветривании помещения.
- Вода для полива должна быть мягкой, не содержащей извести. Подойдет дождевая, кипяченая и отстоянная в течение суток. Зимой в прохладных помещениях поливают реже. Излишняя сырость тоже вредна для папоротников.
- Большинство папоротников выращивают в слабокислой почве, состоящей из смеси равных частей компостной (лучше листовой) земли, торфа и крупного речного песка. Иногда к этой смеси добавляют мох сфагнум, хвойную кору и древесный уголь. Разным видам нужны несколько различные почвенные смеси, например, нефролепису можно добавлять в почву дерновой земли, а платицериуму или асплениуму – нет, для этих папоротников можно использовать почвенную смесь для орхидей.

- Зимой папоротники почти не растут, весной с появлением новых листьев, их удобряют слабым раствором органических и минеральных удобрений.
- Пересаживают по мере необходимости весной после того, как появится первый рост. Посадку производят так, чтобы корневая шейка папоротника находилась над землей. Листья и корни хрупкие почти в всех видов, пересаживать осторожно.
- Размножают папоротники делением куста при пересадке. Некоторые виды папоротников дают новые растения на концах укореняющихся побегов (например, Нефролепис возвышенный, Асплениум луковиценосный).
- Покупать папоротники лучше весной, приобретенные осенью и зимой растения чаще всего не могут адаптироваться в домашних условиях и погибают.

Правильный уход за папоротником в домашних условиях обеспечит растение всем необходимым для нормального развития, а оно порадует своим красивым и здоровым видом.
- < Назад
- Вперёд >
Папоротники для сада: ТОП-17 лучших садовых папоротников
Папоротники очень распространены в дикой природе, но относительно редко встречаются в домашнем пейзаже. Это нечестно, потому что эти изящные, долгоживущие многолетники такие разнообразные! Они распространены в широком диапазоне цветов, форм и размеров. Более 20 000 известных видов папоротников растут во всем мире. Многие виды процветают как на открытом воздухе, так и в качестве комнатных растений. Кроме того, папоротники невероятно легко растут и редко поражаются болезням или вредителями.
Это нечестно, потому что эти изящные, долгоживущие многолетники такие разнообразные! Они распространены в широком диапазоне цветов, форм и размеров. Более 20 000 известных видов папоротников растут во всем мире. Многие виды процветают как на открытом воздухе, так и в качестве комнатных растений. Кроме того, папоротники невероятно легко растут и редко поражаются болезням или вредителями.
Образ папоротника в саду
Папоротники создают образ первозданной природы. В саду они представляют ощущение спокойствия, гармонии и постоянства. Папоротники отлично дополняют любой теневой участок сада.
Если ваш двор затенен в большей его части, это создает некую сложность поддержания красивого газона. Подумайте о замене солнцелюбивых растений папоротниками. Они являются отличным грунтовым покрытием и требуют гораздо меньше внимания, чем даже трава.
Эти реликтовые растения представлены как грозными исполинами, так и в крохотном, удивительно изящном и кружевном исполнении. Каждый вид папоротника наделен уникальной индивидуальностью и выделяется на фоне садовых растений.
Каждый вид папоротника наделен уникальной индивидуальностью и выделяется на фоне садовых растений.
Наиболее декоративные виды
Орляк обыкновенный (Pteridium aquilinum) — широко распространенный папоротник, произрастает во всем мире в умеренных и тропических регионах. Имеет огромные стебли, с возрастом достигают двухметровой высоты. Листья используются в качестве корма, их готовят как овощи и добавляют в супы в некоторых частях Азии.
Растение обычно разделяют на 5-12 подвидов, хотя некоторые ботаники классифицируют большинство разновидностей, как отдельные виды, что является спорным вопросом среди таксономистов. Корни широко расползаются под землей. Отдельные корневища, что было задокументировано, достигают до 400 метров в длину, что делает его одним из крупнейших растений в мире. Листья, несмотря на то, что умирают осенью, часто остаются зелеными в течение всей зимы. Миниатюрные споры невероятно легкие, что позволило растению распространиться почти по всему миру.
Орляк — агрессивный колонизатор открытого грунта и охотно вторгается в пастбища и поля. После укоренения глубокого корневища, его почти невозможно искоренить. Вид считается одним из худших сорняков, и в некоторых странах числится как инвазивный вид.
Стагхорн — принадлежит к семейству платицериум. Это популярный вид папоротников, у которых уникальная особенность листьев. Хотя он обычно растет на коре деревьев в Азии, Африке и Австралии, он является отличным комнатным растением, если он посажен в почве с хорошим дренажем.
Вид имеет два набора листьев. Зеленые листья плодородны, напоминают оленьи рога, имеют споры на нижней стороне и вырастают до 15 см в длину. Коричневые бесплодные листья растут отдельно от зеленых, они короткие, плоские и округлые. Если вы хотите выращивать их в помещении, убедитесь, что этому папоротнику достаточно сырости и богатой гумусом земли.
Бостон (Nephrolepis exaltata) — этот вид папоротника обычно выращивают в висячем кашпо на крыльце.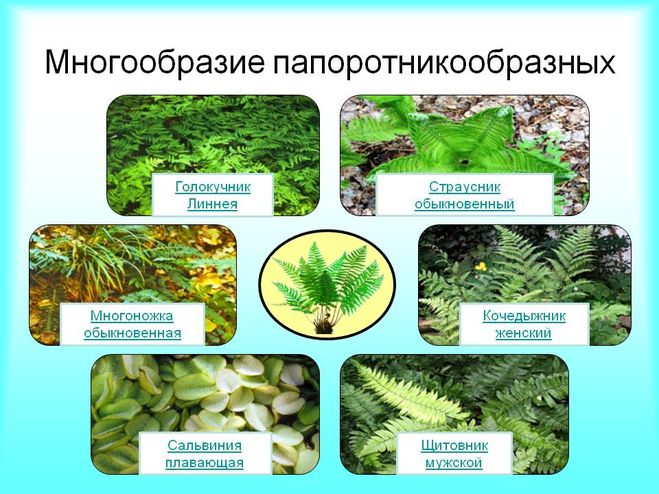 Выделяется темно-зелеными листьями и изогнутыми листьями-перьями. Бостонский папоротник не выдерживает низкой температуры, поэтому держите его в тепле.
Выделяется темно-зелеными листьями и изогнутыми листьями-перьями. Бостонский папоротник не выдерживает низкой температуры, поэтому держите его в тепле.
Холи — маленький папоротник, который имеет узкие глянцевые листья с двойным рядом жестких щетинковых долей. Произрастает главным образом в горных районах как Евразии, так и Северной Америки.
Он предпочитает влажную, хорошо дренированную почву и яркий или рассеянный свет.
Призрачный папоротник — этот прекрасный папоротник является гибридом между леди-папоротником и японским окрашенным папоротником. Растения имеют вертикальный стебель и серебристо-серые листья. Этому виду необходимо обеспечить укрытие от ветра. Призрачные папоротники не будут терпеть засуху, поэтому важно выбрать место посадки, где почва постоянно влажная в течение всего сезона. Растения растут относительно медленно, но в конечном итоге могут распространиться на 0,6 – 0,9 см. Высота от 30 40 см.
Страусник — этот вид папоротника весьма распространен в ландшафте. У него обычная тенденция роста, и к концу лета часто достигает 1,2- 1,5 м в высоту, от 1 до 1,5 м в ширину. Страусиные папоротники производят два вида листьев. Декоративные бесплодные листья длинные и кружевные, как страусиный шлейф. Они выходят из основания растения ранней весной, свернувшись, и медленно разворачиваются до их полной 1,5 м длины. Плодородные листья появляются в середине лета в центре скопления. Они растут примерно до метра и обычно сохраняются зимой.
У него обычная тенденция роста, и к концу лета часто достигает 1,2- 1,5 м в высоту, от 1 до 1,5 м в ширину. Страусиные папоротники производят два вида листьев. Декоративные бесплодные листья длинные и кружевные, как страусиный шлейф. Они выходят из основания растения ранней весной, свернувшись, и медленно разворачиваются до их полной 1,5 м длины. Плодородные листья появляются в середине лета в центре скопления. Они растут примерно до метра и обычно сохраняются зимой.
Страусник — предпочитает среднюю влажность, слегка кислые почвы и полную или частичную тень. В идеальных условиях образуют крупные плотные колонии. Поэтому важно дать им достаточно места для распространения.
Кожаный папоротник — этот вид является родным для восточной части США, и известен как щитовник. Он достигает высоты 2 метров. Имеет темно-зеленые до сине-зеленых листья шириной от 15 до 30 см и текстуру кожи. Щитовник выращивают во влажной почве и защищенном от солнца и сухих ветров месте. Этот компактный папоротник — отличный выбор для любого тенистого сада.
Этот компактный папоротник — отличный выбор для любого тенистого сада.
Леди в красном (Athyrium filix-femina Lady in Red) — являются родными для восточной и центральной части США. Этот вид выделяется ярко-красным стеблем. Глубокорезные светло-зеленые листья имеют ширину от 20 до 30 см и длину 0,6-0,9 см. Глянцевые красные стебли усиливаются в цвете, когда растения созревают. Растет до 2 м высоты и до 1,5 ширины. «Леди в красном» — отличное почвопокровное растение и хороший компаньон для других многолетников. Листва — отличный наполнитель для цветочных композиций.
Рождественский — легко адаптируемый папоротник, который предпочитает расти в частичной тени и будет терпеть как влажные, так и сухие почвы. Глянцевые, темно-зеленые ветви рождественского папоротника имеют длину 20 см и ширину от 2,5 до 7,5 см. Зимой они лежат на земле и сохраняют свой зеленый цвет.
Сколопендриум — маленький вертикальный папоротник, которому нужна влажная почва и укрытие от полуденного солнца. Язычковые листья нуждаются в тщательном укрытии, потому что они легко выгорают и становятся желтоватыми. Края листьев выглядят так, будто они «гофрированы». Найдите подходящее место, и он будет сиять контрастом необычных листьев.
Язычковые листья нуждаются в тщательном укрытии, потому что они легко выгорают и становятся желтоватыми. Края листьев выглядят так, будто они «гофрированы». Найдите подходящее место, и он будет сиять контрастом необычных листьев.
Кристас— король среди папоротников. Этот вертикальный изящный папоротник достигает 90 см. Темно-зеленые гребни листьев на верхушке, молодые по бокам имеют булавки, что добавляет им вес, так что концы листьев опускаются к земле.
Корнуоллский— воздушный ажурный папоротник нуждается во влаге. Светло-зеленые кружевные листья очень изящны.
Японский (Athyrium nipponicum)— это маленькое растение, которое растет до 80 см и имеет великолепный серебристо-серый цвет, с оттенком бордо. Этот вид очень эффектный.
Майденхара — имеет тонкие листья и часто выращивается как комнатное растение. Это великолепное вечнозеленое растение растет всего 0,4-0,6 см в высоту и любит влагу и тень.
Австралийский — этот папоротник можно выращивать в большом контейнере в качестве дерева патио и зимой в закрытом помещении. Действительно, он быстро растет до 3 м в контейнере, 6 м в земле. Этот вид вечнозеленого папоротника требует частичного оттенка и большого количества влаги. Он добавляет экзотики в любое пространство!
Аспарагус — имеет прекрасные игольчатые листья, которые раздражают кожу. Они хорошо развиваются на ярком солнце и часто размножаются так хорошо, что захватывают целые сады, поэтому держите их под контролем и часто обрезайте.
Синамон (Osmundastrum cinnamomeum) — этот папоротник в дикой природе часто растет вдоль ручьев, поэтому для посадки в саду требуется много влаги. Достигает около 1,5 м в высоту и имеет два вида листьев. Ярко-зеленые листья — бесплодные, а плодородные имеют глубокий цвет корицы.
Лучшие партнеры
Для того чтобы папоротники совершенно вписались в пейзаж садовых многолетников, им необходимы достойные партнеры. Традиционно композицию папоротникам составят декоративно-лиственные растения, такие как бадан, роджерсии, киренгешомы веерные. Используйте для фоновых насаждений или комбинируйте любящими тень хостами, астильбами, дицентрами и калладией.
Традиционно композицию папоротникам составят декоративно-лиственные растения, такие как бадан, роджерсии, киренгешомы веерные. Используйте для фоновых насаждений или комбинируйте любящими тень хостами, астильбами, дицентрами и калладией.
Присоединяйтесь к нашей группе в Facebook
Папоротникообразные
Вы здесь: Главная – Папоротникообразные
В данном разделе представлены папоротниковидные (или папоротники) занесённые в Красную Книгу России.
Красная книга содержит сведения о распространении и мерах охраны редких и находящихся под угрозой исчезновения видов папоротников, произрастающих на территории Российской Федерации, сведения о численности, особенностях экологии и пр.
Папоротниковидные (или папоротники) относят к числу наиболее древних групп высших растений, в настоящее время насчитывается около 300 родов и более 10 ООО видов папоротников.
Папоротники занесённые в Красную книгуПапоротники распространены очень широко, фактически по всему земному шару, и встречаются в самых различных местообитаниях, начиная с пустынь и кончая болотами, озерами, рисовыми полями и солоноватыми водами. По своим размерам папоротники варьируют от тропических древовидных форм (иногда достигающих высоты 25 м, а диаметра ствола 50 см) до крошечных растеньиц длиной всего лишь в несколько миллиметров.
По своим размерам папоротники варьируют от тропических древовидных форм (иногда достигающих высоты 25 м, а диаметра ствола 50 см) до крошечных растеньиц длиной всего лишь в несколько миллиметров.
Несмотря на видовое многообразие и многочисленность папоротников, многие виды находятся на грани исчезновения или их уже нет.
Материалы по теме
Костенец черный
Царство: Растения Отдел: Папоротниковидные Класс: Папоротниковые Порядок: Многоножковые Семейство: Костенцовые Род: Костенец Вид: Костенец чёрный Костенец черный - Asplenium adiantum-nigrum L Категория и статус: 3 б - редкий вид.
AOF | 06.06.2017 12:18:30 | Папоротникообразные
ПодробнееКостенец дагестанский
Царство: Растения Отдел: Папоротниковидные Класс: Папоротниковые Порядок: Многоножковые Семейство: Костенцовые Род: Костенец Вид: Костенец дагестанский Костенец дагестанский - Asplenium daghestanicum Christ Категория и статус: 3 а - редкий вид.
 Узколокальный эндемик Дагестана
Узколокальный эндемик ДагестанаAOF | 06.06.2017 12:17:25 | Папоротникообразные
ПодробнееКостенец алтайский
Царство: Растения Отдел: Папоротниковидные Класс: Папоротниковые Порядок: Многоножковые Семейство: Костенцовые Род: Костенец Вид: Костенец алтайский Костенец алтайский - Asplenium altajense (Kom.) Grub. Категория и статус: 3 г - редкий вид, имеющий в России северную границу ареала.
AOF | 06.06.2017 12:16:28 | Папоротникообразные
ПодробнееДалее…
Мосприрода проведет экспедицию / Новости города / Сайт Москвы
Экспедицию «В поисках цветущего папоротника» специалисты Мосприроды проведут в природно-историческом парке «Кузьминки-Люблино» 2 августа в 14:00. Все желающие могут стать исследователями и внести свой вклад в поиск редких растений.
Все желающие могут стать исследователями и внести свой вклад в поиск редких растений.
Экспедиция приурочена к празднику Ильин день – церковный день памяти пророка Илии и народный праздник у славян. Илья в славянской народной традиции – повелитель грома, небесного огня, дождя, покровитель урожая и плодородия. Ильин день считался календарной границей лета, когда в природе появлялись первые признаки осени. При этом во второй половине лета травы набирают свой максимальный размер и концентрацию активных веществ, многие из которых обладают целительным действием, поэтому можно было отправляться на поиск диких лекарственных растений, одним из которых был мифический цветок папоротника.
В Кузьминском парке произрастают редкие виды папоротников, занесенные в Красную книгу Москвы – телиптерис болотный и гроздовник полулунный. Но, безусловно, все виды папоротников, как и редкие, так и обычные, нельзя рвать на особо охраняемой природной территории, ведь, как и все другие растения, они являются важной частью природного биотопа. Кроме того, хотя цветков у папоротника не бывает, возможно, не зря им приписывали в старину волшебные свойства, и в будущем будут открыты новые применения этих древних растений.
Кроме того, хотя цветков у папоротника не бывает, возможно, не зря им приписывали в старину волшебные свойства, и в будущем будут открыты новые применения этих древних растений.
Участники похода будут искать не только папоротники, но и другие редкие растения. Также экологи планируют встретить кузнечиков, стрекоз и краснокнижных обитателей природной территории. Редкие животные и растения наверняка попадутся на маршруте, ведь в городе зафиксировано 569 видов различной флоры и фауны, которые бережно охраняются и восстанавливаются.
2 августа ждем вас по адресу: ЮВАО, ул. Кузьминская, д. 10, с. 1. Для записи: 8 (495) 377-35-93.
Мероприятие 12 +.
Участие бесплатное.
Напоминаем, что в группах действуют ограничения по количеству человек и соблюдаются меры предосторожности. Кроме того, отправляясь на природу, не забывайте о мерах личной безопасности: необходимо одеться так, чтобы клещи не могли заползти под одежду.
Гастрономические изыски у нас под ногами
Квас из бузины, черемшовая паста и тушенный кавказский папоротник – члены Русского географического общества готовы удивить самых требовательных гурманов.
Походное меню
Идея изучать кухню из дикорастущих растений появилась в ходе длительных экспедиций, в которых участвовали члены Горячеключевского местного отделения Краснодарского регионального отделения Русского географического общества им. А.В. Твердого. В таких походах как правило очень скудный запас пищи, он помещается в рюкзаке, который человек несет на себе. А это значит, что из меню в рационе туриста присутствует только самое необходимое.
Между тем, практически в каждом регионе произрастают растения, которые можно употреблять в пищу – грибы, ягоды, травы. Например, корень лопуха вполне может заменить картошку в супе или ухе, а различные травы боярышник, чабрец, душица, мята и другие – это прекрасный фиточай. Об этом знает практически каждый турист с рюкзаком.
Дикоро́сы — под этим термином учёные понимают дикорастущие, то есть не культивируемые плодовые тела грибов, плоды и семена дикорастущих растений.
Пищевые дикоросы всегда присутствовали в рационе наших предков: крапива, лебеда, щавель, сныть, черемша, цикорий, лопух. Для рациона сибиряков и жителей севера европейской части России эти природные дары традиционны и сегодня. Во время блокады Ленинграда употребляли около 40 видов дикорастущих трав, добавляя их в обычную еду или используя как самостоятельные овощи.
Многие дикорастущие растения по своей пищевой ценности превосходят культивируемые. Так, в крапиве витамина С содержится в 8 раз больше чем в капусте кольраби, содержание каротина в крапиве — в полтора раза выше, чем в петрушке, а по содержанию протеинов лебеда не уступает шпинату. Нередко зелень съедобных дикоросов обладает высокой лечебной активностью, благодаря чему находит свое применение в фитотерапии и народной медицине.

Настоящие деликатесы зачастую растут прямо у нас под ногами. Например, на Кавказе в мае распускается папоротник, некоторые виды которого не только можно употреблять в пищу, но и приготовить из них очень изысканные блюда.
Сезонные предложения
Начало экспедиционного сезона – прекрасное время, чтобы попробовать яства из папоротника орляка. Папоротник считается одним из древнейших растений на земле. Он рос уже 405 миллионов лет назад, его ели динозавры и первые млекопитающие. Папоротник-орляк, благодаря своим отличным вкусовым качествам и полезным питательным свойствам, издревле широко использовался в кулинарии разных народов мира и в народной медицине.
Его отваривают, жарят, тушат, маринуют, и засаливают. Ростки орляка собирают в самом начале сезона, когда папоротник еще не раскрылся, а только выпустил свои первые побеги. При этом походные повара проявляют очень бережное отношение к молодому растению – с каждой розетки можно срывать только 2-3 побега, не больше, чтобы не нарушить его созревание. Если папоротник употребляют свежим, то его нужно отварить 7–8 минут в соленой воде, потом промыть и дать воде стечь на дуршлаге или сите. После этого его можно жарить, тушить, мариновать. Из папаротника готовят салаты, варят супы и вторые блюда, делают приправу, а также солят и маринуют. По вкусу орляк напоминает грибы, иногда похож на спаржу.
Из папаротника готовят салаты, варят супы и вторые блюда, делают приправу, а также солят и маринуют. По вкусу орляк напоминает грибы, иногда похож на спаржу.
В этот же период можно собрать и черемшу, из которой готовится вкусный соус. Рецепт прост: перемолоть листья черемши в блендере, добавить оливковое масло и немного соли. Получится “черемшовая паста” ярко-зеленого цвета. Ее можно хранить в морозильнике, а зимой использовать в качестве приправы для блюд или соуса в сочетании с майонезом.
В конце мая путешественники собирают цветы бузины древовидной. Из этих белых соцветий можно приготовить ароматный квас. Необходимо 15 соцветий бузины залить водой (не кипяченой и не фильтрованной), добавить нарезанный мелко лимон, сахар (на 4 литра воды добавляем примерно 400 гр сахара) и оставить примерно на неделю. Через семь дней у вас получится прекрасный квас с неповторимым ароматом.
Поход на выживание
Чтобы превратить обычный поход в настоящее приключение, достаточно отправиться в путь без еды. В такой “поход на выживание” осенью 2016 года отправились председатель Горячеключевского местного отделения Общества им. А.В. Твердого Александр Емельяненко, члены РГО Роман Волощенко и Владислав Карпинский. Автором идеи и руководителем похода стал член совета Горячеключевского местного отделения Общества Сергей Ткач. Основной целью туристов было изучение возможностей добычи провианта в полевых условиях в осенний период.
В такой “поход на выживание” осенью 2016 года отправились председатель Горячеключевского местного отделения Общества им. А.В. Твердого Александр Емельяненко, члены РГО Роман Волощенко и Владислав Карпинский. Автором идеи и руководителем похода стал член совета Горячеключевского местного отделения Общества Сергей Ткач. Основной целью туристов было изучение возможностей добычи провианта в полевых условиях в осенний период.
Помимо традиционного туристского снаряжения путешественники положили в свои рюкзаки минимальный набор вспомогательных продуктов – муку, соль, соевый соус и сахар. Остальные съестные припасы участникам экспедиции предстояло отыскать уже в пути.
На маршрут группа вышла в Горячем ключе, далее проследовала через урочище Поднависла, вдоль долины реки Чепси до хребта Хазарова. После чего путешественники спустились в долины рек Бурхан и Шапсухо и уже оттуда вышли на конечную точку – поселок Лермонтово. Общая протяженность пешего маршрута составила более семидесяти километров.
В течении шести дней основу меню путешественников составляли грибы, обилие которых в осеннем лесу могло бы порадовать даже искушенных гурманов. Улов участников похода также отличался разнообразием. В числе рыбных блюд – форель, голавль и бычки в кляре. Не обошлось в походе и без деликатесов. Корень лопуха жареный в соевом соусе и варенье из груши-дички также вошли в рацион туристов. Отличались вкусовой палитрой и напитки: чаи из шиповника и пихты с лимоном обладают тонким ароматом и повышают иммунитет.
Подобный формат “похода на выживание” имеет ряд характерных особенностей. Он наиболее приемлем для небольших групп – 4-5 человек. В таком составе участники могут распределить усилия по добыче еды и не остаться голодными. Меняется и структура похода. Время, потраченное на поиски и приготовление пищи, существенно сокращает преодолеваемые расстояния.
“О выживании и речи не могло быть, – отметил руководитель похода Сергей Ткач, – в наших лесах так много съедобных растений, что еще нужно очень постараться “умереть с голоду” или хотя бы всерьез проголодаться, тем более осенью”.
Традиции дегустаций
Кухня из дикоросов – визитная карточка многих фестивалей, мастер-классов и научно-популярных лекториев Краснодарского регионального отделения Русского географического общества. В планах у членов РГО совершить гастрономические путешествия в лес в разные времена года, чтобы объективно оценить возможность выживания в условиях похода без запаса провизии.
На разных мероприятиях РГО, гости с большим удовольствием уже отведали тушенный кавказский папоротник, соусы из щавеля и черемши, а также ткемали из тёрна. Традиционно дегустации сопровождаются познавательным рассказом о том, какие виды папоротников произрастают в наших лесах, какие из них съедобные, и как правильно заготавливать их побеги.
Осенью мастер походной кухни Сергей Ткач угощал участников интеллектуальной площадки Горячеключевкого местного отделения Общества “Открытое пространство” гренками с каштановым пюре. Для многих было удивительно, что каштаны можно употреблять не только в жаренном или печеном виде, но и готовить из них суп или пюре, в лучших традициях французской кухни. Каштаны – настоящая находка для любителей диетического стола.
Каштаны – настоящая находка для любителей диетического стола.
При относительно невысоком содержании жиров, в них содержится масса витаминов и легких углеводов. А на фестивале “Вкус лета”, который открыл в этом году курортный сезон в Горячем Ключе, Сергей Ткач давал гостям попробовать тушеный папоротник в сочетании с рисом, приправленный соусом из черемши.
Ксения Руденко
Dryopteris: Древесный папоротник
Dryopteris – самый многочисленный род папоротников, насчитывающий более 200 видов и множество гибридов. Это привлекательные декоративные папоротники с прямостоячими изогнутыми ветвями, которые отлично смотрятся в смешанных границах с другими многолетниками.
Многие виды вечнозеленые. Когда ножки разворачиваются, многие из Dryopteris имеют привлекательные чешуйки на кончиках, а также вдоль основания ножек у зрелых листьев.
Здесь, на северо-западе Тихого океана, почти всем знакомо появление папоротников в лесах. Большие папоротники-мечи прижимаются к земле под высокими кедрами и дугласовыми елями, в то время как папоротники девичьей шерсти изящно трепещут, цепляясь за камни у водопада. Оленьи папоротники будут сюрпризом на уровне глаз, растущие на узловатой коре старого клена. Древесные папоротники встречаются в лесах, полях и влажных районах Северного полушария, многие из которых встречаются в Восточной Азии и Гималаях.
Большие папоротники-мечи прижимаются к земле под высокими кедрами и дугласовыми елями, в то время как папоротники девичьей шерсти изящно трепещут, цепляясь за камни у водопада. Оленьи папоротники будут сюрпризом на уровне глаз, растущие на узловатой коре старого клена. Древесные папоротники встречаются в лесах, полях и влажных районах Северного полушария, многие из которых встречаются в Восточной Азии и Гималаях.
Папоротники также являются полезными садовыми растениями, особенно в тенистом или лесном саду. Многочисленные разновидности древесного папоротника добавляют мягкости и текстуры. Они могут быть растением-наполнителем в тенистом саду, или, поскольку многие Dryopteris вечнозеленые, они могут быть ярким растением. Особенно хорошо они сочетаются с декоративными травами hostas и Farfugium .
Многие Dryopteris имеют красочные новообразования оттенков бронзы или красного.Эта очаровательная особенность делает их идеальными участниками конкурса контейнеров и подвесных корзин. Несколько деревянных папоротников в подвесной корзине могут воссоздать классические корзины из бостонских папоротников, которые висят на балконах Французского квартала в Новом Орлеане. Дополнительным бонусом является то, что древесные папоротники выносливы в нашем климате, и если вы выберете вечнозеленый, он будет хорошо смотреться круглый год.
Несколько деревянных папоротников в подвесной корзине могут воссоздать классические корзины из бостонских папоротников, которые висят на балконах Французского квартала в Новом Орлеане. Дополнительным бонусом является то, что древесные папоротники выносливы в нашем климате, и если вы выберете вечнозеленый, он будет хорошо смотреться круглый год.
Dryopteris свободно гибридизируются в дикой природе, что иногда затрудняет их идентификацию, но также дает множество различных растений и особенностей, которые делают их хорошими декоративными растениями.Все Dryopteris – прочные, сильнорослые садовые растения с небольшими проблемами, что делает их простыми и универсальными. Лучше всего то, что они практически не требуют обслуживания.
Разновидностей в питомнике Портленда включают:
Dryopteris cycadina ( Dryopteris atrata ) Shaggy Shield Папоротник Вечнозеленое растение с длинными темно-зелеными листьями и зубчатыми краями листочков.
Листья имеют дугообразную форму с черными стеблями, около 2–3 футов в высоту и 18–24 дюйма в ширину.
Черные стебли на новых приростах.
Зоны 5-8.
Dryopteris erythrosora ‘Brilliance’: Блестящий осенний папоротникСимпатичный вечнозеленый папоротник с новой бронзовой листвой, ослепительно контрастирующей с темно-зелеными более старыми листьями.
Может выдерживать утреннее солнце или находиться в тени весь день, высота и ширина 18–24 дюйма.
Отличное акцентное растение в теневом контейнере или зимнем цветном горшке.
Зоны устойчивости 5-9.
Dryopteris felix-mas: Папоротник самец Вечнозеленый или полувечнозеленый папоротник, отлично подходит для массовых посадок, а также для лесных садов или бордюров.
Легкие в выращивании, сильнорослые виды
Развивается на частично затененных влажных участках высотой 2-3 фута и шириной 2 фута.
Зоны 4-8.
Разновидности включают:
- ‘Barnseii’ – особенно вертикальная форма
- ‘Linearis Polydactyla’ – тонко остриженные, очень кружевные листья
- ‘Петрушка’ – листья экстра-взъерошенные
Evergreen с великолепными бронзовыми, розовыми и золотыми новообразованиями.
Вырастает 24-30 дюймов в высоту и ширину во влажной тени или частично затененной местности.
Выносливость в зонах 6-9
Dryopteris tokyoensis: Токийский лесной папоротник Листопадные, яблочно-зеленые листья вырастают 18-36 дюймов в высоту.
Чрезвычайно прямые листья создают сильное вертикальное присутствие
Полная или частичная тень.
Зоны 5-8
границ | Функциональное разнообразие папоротников обусловлено скорее видовым богатством, чем ограничениями окружающей среды
Введение
Исследования функционального разнообразия все чаще используются для понимания процессов сборки сообществ (Petchey and Gaston, 2006; Mouchet et al., 2010; Lamanna et al., 2014; Tanaka and Sato, 2015; Pigot et al., 2016). Функциональное разнообразие измеряет то, что делают организмы, а не их таксономическую идентичность, и, таким образом, учитывает функциональную комплементарность и избыточность сопутствующих видов (Diaz and Cabido, 2001; Petchey and Gaston, 2006).Функциональное разнообразие можно понять с точки зрения концепции ниши Хатчинсона (Hutchinson, 1957), то есть путем определения распределения видов в функциональном пространстве, оси которого являются чертами, прямо или косвенно связанными с функцией (Rosenfeld, 2002). Этот подход критически зависит от выбора признаков, и, следовательно, осознанный выбор, основанный на экологической функции, имеет жизненно важное значение.
Этот подход критически зависит от выбора признаков, и, следовательно, осознанный выбор, основанный на экологической функции, имеет жизненно важное значение.
Функциональные признаки – это морфологические, анатомические, физиологические, репродуктивные или поведенческие характеристики, которые влияют на то, как вид адаптируется к своей абиотической и биотической среде и взаимодействует с ней.Анатомические и физиологические признаки, описываемые экономическим спектром листьев, часто изучаются на растениях (Bruelheide et al., 2018), хотя их трудно отобрать в крупномасштабных исследованиях. Среди папоротников морфологические признаки, такие как жизненная форма и размер растения, широко используются для описания функционального разнообразия (Kluge and Kessler, 2007; Tanaka and Sato, 2015; Carvajal-Hernández et al., 2018; Nitta et al., 2020).
Кроме того, поскольку функциональность признаков имеет разные аспекты, необходимо использовать дополнительные индексы, чтобы лучше понять, какие правила управляют процессами сборки (Mouchet et al. , 2010). Например, объем признака представляет собой объем пространства признака, покрытого сборкой. Насколько равномерно распределены виды в пространстве признаков, будет напрямую зависеть от того, насколько эффективно сообщество использует весь диапазон доступных ресурсов. Наконец, плотность относится к тому, как упакованы виды внутри этого объема, что указывает на степень дифференциации ниши и конкуренции за ресурсы.
, 2010). Например, объем признака представляет собой объем пространства признака, покрытого сборкой. Насколько равномерно распределены виды в пространстве признаков, будет напрямую зависеть от того, насколько эффективно сообщество использует весь диапазон доступных ресурсов. Наконец, плотность относится к тому, как упакованы виды внутри этого объема, что указывает на степень дифференциации ниши и конкуренции за ресурсы.
Простая концептуальная модель экологической сборки утверждает, что как только вид попадает на место, он сначала проходит через абиотический фильтр (т.д., фильтрация окружающей среды), которая определяет, какие морфологические и физиологические признаки позволяют виду сохраняться в заданном наборе экологических условий (Weiher and Keddy, 1995). В экстремальных условиях может быть выбрано ограниченное количество характеристик, позволяющих сохранять устойчивость в этих конкретных условиях. В этих сценариях сосуществующие виды будут сближаться функционально (Grime, 2006), приводя к более плотно упакованным областям по сравнению с другими, просто пустыми в морфопространстве, и уменьшая равномерность распределения признаков. Более того, увеличение упаковки видов с большей вероятностью приведет к уменьшению объема, чем ожидалось случайно. Затем виды проходят через биотический фильтр конкуренции сосуществующих видов, содействия или общих врагов. В частности, конкуренция может привести к расхождению в функциональной стратегии (ограничивающая теория сходства; MacArthur and Levins, 1967), что приведет к увеличению объема признаков с увеличением видового богатства, что приведет к снижению плотности и более равномерному распределению видов. Содействие и общие враги могут приводить к сильно различающимся эффектам на видовые черты в пространстве черт с закономерностями, которые трудно предсказать (Garzon-Lopez et al., 2015; Мишале и Пуньер, 2016).
Более того, увеличение упаковки видов с большей вероятностью приведет к уменьшению объема, чем ожидалось случайно. Затем виды проходят через биотический фильтр конкуренции сосуществующих видов, содействия или общих врагов. В частности, конкуренция может привести к расхождению в функциональной стратегии (ограничивающая теория сходства; MacArthur and Levins, 1967), что приведет к увеличению объема признаков с увеличением видового богатства, что приведет к снижению плотности и более равномерному распределению видов. Содействие и общие враги могут приводить к сильно различающимся эффектам на видовые черты в пространстве черт с закономерностями, которые трудно предсказать (Garzon-Lopez et al., 2015; Мишале и Пуньер, 2016).
В конечном итоге, изменения в объеме признаков отражают два взаимодополняющих процесса: расширение функциональной ниши и упаковка ниши (Swenson and Weiser, 2014). Когда виды выбираются случайным образом из пула видов при низком уровне видового богатства, объем признаков, вероятно, будет увеличиваться по мере добавления новых видов, потому что они имеют комбинации признаков за пределами ранее занятого пространства ниши. На более высоких уровнях видового богатства дополнительные виды с большей вероятностью попадут в уже занятое пространство признаков, увеличивая упаковку видов (Pigot et al., 2016). Процессы расширения и упаковки функциональных ниш широко изучались на животных (Ricklefs and O’Rourke, 1975; Ricklefs and Cox, 1977). Например, у птиц Анд высокое видовое богатство в основном связано с функциональной упаковкой ниш, что дает четкую картину насыщения функционального разнообразия (Pigot et al., 2016). Паттерны расширения и упаковки функциональных ниш менее изучены у растений, но Свенсон и Вейзер (2014) обнаружили увеличение объема и упаковки ниш с богатством видов в сообществах деревьев восточной части Северной Америки без насыщения функционального разнообразия.
На более высоких уровнях видового богатства дополнительные виды с большей вероятностью попадут в уже занятое пространство признаков, увеличивая упаковку видов (Pigot et al., 2016). Процессы расширения и упаковки функциональных ниш широко изучались на животных (Ricklefs and O’Rourke, 1975; Ricklefs and Cox, 1977). Например, у птиц Анд высокое видовое богатство в основном связано с функциональной упаковкой ниш, что дает четкую картину насыщения функционального разнообразия (Pigot et al., 2016). Паттерны расширения и упаковки функциональных ниш менее изучены у растений, но Свенсон и Вейзер (2014) обнаружили увеличение объема и упаковки ниш с богатством видов в сообществах деревьев восточной части Северной Америки без насыщения функционального разнообразия.
Папоротники и ликофиты (далее для простоты называемые папоротниками) являются полезной исследовательской группой для оценки структур биоразнообразия из-за их глобального распространения, составляющих до 70% видового богатства местной тропической флоры, а также из-за того, что они имеют достаточно высокий, но управляемый вид. богатство (Kreft et al., 2010). В частности, диспергирование спор снижает их зависимость от биотических векторов распространения, в то же время позволяя им легко колонизировать подходящие среды обитания, так что мешающие эффекты ограничения распространения на процессы сборки снижаются (Wolf et al., 2001; Karger et al., 2014). Предыдущие исследования функционального разнообразия папоротников вдоль высотных градиентов показали, что разнообразие морфологических признаков отражает структуру видового богатства. В Коста-Рике и Мексике видовое богатство и разнообразие признаков демонстрируют горбовидную структуру с максимальными значениями на средней высоте, в Японии – монотонный спад, а на острове Муреа во Французской Полинезии – линейное увеличение эпифитов (Kluge and Kessler, 2007; Tanaka, Sato, 2015; Carvajal-Hernández et al., 2018; Nitta et al., 2020). Используя нулевую модель для исключения влияния видового богатства, Клюге и Кесслер (2007) обнаружили, что для эпифитных папоротников в Коста-Рике средние возвышения имели более высокое, чем ожидалось, разнообразие признаков, тогда как верхние и нижние возвышения сохраняли меньшее разнообразие.
богатство (Kreft et al., 2010). В частности, диспергирование спор снижает их зависимость от биотических векторов распространения, в то же время позволяя им легко колонизировать подходящие среды обитания, так что мешающие эффекты ограничения распространения на процессы сборки снижаются (Wolf et al., 2001; Karger et al., 2014). Предыдущие исследования функционального разнообразия папоротников вдоль высотных градиентов показали, что разнообразие морфологических признаков отражает структуру видового богатства. В Коста-Рике и Мексике видовое богатство и разнообразие признаков демонстрируют горбовидную структуру с максимальными значениями на средней высоте, в Японии – монотонный спад, а на острове Муреа во Французской Полинезии – линейное увеличение эпифитов (Kluge and Kessler, 2007; Tanaka, Sato, 2015; Carvajal-Hernández et al., 2018; Nitta et al., 2020). Используя нулевую модель для исключения влияния видового богатства, Клюге и Кесслер (2007) обнаружили, что для эпифитных папоротников в Коста-Рике средние возвышения имели более высокое, чем ожидалось, разнообразие признаков, тогда как верхние и нижние возвышения сохраняли меньшее разнообразие. Они интерпретировали это как отражение фильтрации окружающей среды на крайних уровнях градиента и биотической фильтрации в середине. Аналогичные выводы были сделаны для возвышенностей в Японии (Tanaka, Sato, 2015). Однако эти исследования ограничиваются отдельными трансектами, а также из-за особенностей видового богатства и изменчивости окружающей среды (температуры, осадки и т. Д.).) тесно взаимосвязаны, сложно по-настоящему изолировать влияние видового богатства и окружающей среды друг от друга. Напротив, если сравнивать несколько трансект, можно обнаружить разные уровни видового богатства на одинаковых высотах и в одинаковых экологических условиях, что потенциально позволяет разделить их соответствующее влияние на состав сообществ.
Они интерпретировали это как отражение фильтрации окружающей среды на крайних уровнях градиента и биотической фильтрации в середине. Аналогичные выводы были сделаны для возвышенностей в Японии (Tanaka, Sato, 2015). Однако эти исследования ограничиваются отдельными трансектами, а также из-за особенностей видового богатства и изменчивости окружающей среды (температуры, осадки и т. Д.).) тесно взаимосвязаны, сложно по-настоящему изолировать влияние видового богатства и окружающей среды друг от друга. Напротив, если сравнивать несколько трансект, можно обнаружить разные уровни видового богатства на одинаковых высотах и в одинаковых экологических условиях, что потенциально позволяет разделить их соответствующее влияние на состав сообществ.
В настоящем исследовании мы сравнили четыре тропических высотных разреза с различными характеристиками окружающей среды и видовым богатством, в том числе некоторые из самых разнообразных ассоциаций папоротников в мире в Эквадоре и Новой Гвинее, а также гораздо более бедные в Мексике и Уганде. Мы стремились отделить влияние видового богатства и окружающей среды на функциональное разнообразие и оценить относительную важность функционального расширения или упаковки с изменениями видового богатства, оценивая морфологические черты, связанные с приобретением ресурсов. В частности, мы проверили следующие гипотезы:
Мы стремились отделить влияние видового богатства и окружающей среды на функциональное разнообразие и оценить относительную важность функционального расширения или упаковки с изменениями видового богатства, оценивая морфологические черты, связанные с приобретением ресурсов. В частности, мы проверили следующие гипотезы:
h2: Функциональное разнообразие в основном определяется (а) биотической фильтрацией (измеряется как видовое богатство) или (б): фильтрацией окружающей среды.
h3: При низком уровне видового богатства увеличение количества видов приводит к увеличению объема признаков, тогда как при высоких уровнях видового богатства это приводит к увеличению плотности в уже занятом объеме признаков.
Материалы и методы
Выбор трансекта и отбор проб растительности
Полевые исследования проводились в 2009–2014 гг. По перепадам высот в Эквадоре (500–4 050 м), Мексике (300–3 000 м), Папуа-Новой Гвинее (200–3 700 м) и Уганде (700–3200 м), при этом все трансекты, представляющие собой горбовидную связь с высотой (рис. 1). Эти трансекты вместе представляют три основных тропических биогеографических района (Америка, Африка и Азия-Океания) с разным уровнем регионального видового богатства (Weigand et al., 2020) и сильно различающийся климат, что позволяет статистически отделить влияние видового богатства от климата. В полевых условиях мы зарегистрировали эпифитные и наземные виды папоротников на 116 площадках размером 20 × 20 м 2 каждый. Участки были размещены в естественных лесах на склонах, избегая нарушенной растительности, а также оврагов, гребней, скал или других местных микропредприятий. Все виды папоротников на каждой трансекте (но не на каждом участке) были собраны и сохранены в гербарии для последующей идентификации и оценки морфологических признаков.Эпифиты были идентифицированы и подсчитаны в бинокль, ища упавшие ветви на земле, лазая по деревьям и обрезая отдельные ветви. Численность каждого вида была определена количественно, но для нашего анализа мы использовали только данные о наличии / отсутствии. Подробное описание полевых методов можно найти в более ранних публикациях (Kessler, Bach, 1999; Kluge, Kessler, 2007; Karger et al., 2014, 2015). Из-за наличия и доступности местообитаний не все высотные разрезы начинались на одной и той же отметке, а участки не располагались на одинаковых отметках.Для анализа мы определили интервалы 500 м, начиная с интервала 0–400 м (таблица 1). Каждый интервал включал четыре участка на каждую трансекту, и все виды, зарегистрированные на этих четырех участках, были объединены в список видов на интервал, который лег в основу анализа. Чтобы оценить, может ли различная полнота выборки повлиять на наши результаты, мы использовали оценку богатства Чао (Chao, 1987) в EstimateS (Colwell, 2013), чтобы оценить общее количество видов в каждом интервале. Расчетное значение богатства составляло 1.На 6–90% (медиана 21%) больше, чем наблюдаемое количество видов (дополнительная таблица S1), что указывает на то, что выборка видов не производилась даже между интервалами. Однако линейный регрессионный анализ наблюдаемого и оцененного количества видов дал R 2 значений 0,84–0,96 для отдельных разрезов и 0,93 в целом (дополнительный рисунок S1), показывая, что неполнота выборки была примерно равномерно распределена по нашему набору данных и не сильно повлияли на наши результаты, основанные на наблюдаемой численности видов.Мы не проводили наши анализы с оценкой количества видов, потому что они часто смещены (Herzog et al., 2002), а также потому, что наши измерения признаков были основаны на фактических данных о видах и не могут быть экстраполированы.
Рисунок 1 . Годовое количество осадков (непрерывные линии) и богатство видов папоротников (столбцы) по четырем тропическим градиентам высот. Общее количество видов разделено на эпифиты (светло-серые), наземные (черные) и виды, которые были обнаружены как эпифиты, так и наземные (темно-серые).Серая полоса на карте мира указывает область между тропиками Козерога и Рака.
Таблица 1 . Количество видов в каждом интервале высот и всего на четырех исследуемых разрезах.
Морфологические признаки
Мы включили морфологические признаки (далее «морфологические признаки»), связанные с использованием ресурсов, то есть потребление света, воды и питательных веществ, поскольку они могут быть напрямую связаны с конкуренцией за ресурсы между видами. Из списка функциональных морфологических признаков папоротников Kluge and Kessler (2007) мы выбрали форму и размер корневища и пластинок, ламинарную текстуру (которую они назвали толщиной) и наличие гидатодов (таблица 2).Мы не использовали признаки, связанные с размножением (например, наличие индусии) или типом и плотностью волосяного покрова. Хотя они, несомненно, функционально значимы, они не связаны напрямую с приобретением ресурсов, и их влияние, если таковое имеется, на конкурентные взаимодействия неясно. Следовательно, если бы мы включили их, мы бы смешали разные адаптивные и выборочные области. Признаки были собраны в основном из соответствующей литературы (например, Holttum, 1991; Stolze and Pacheco, 1994; Verdcourt et al., 1999/2008; Mickel and Smith, 2004), а недостающая информация была получена при изучении гербарных образцов.Поскольку ламинарная текстура демонстрирует внутривидовую изменчивость, чтобы избежать претензий на точность, мы использовали широкое определение состояния характера, как описано в таблице 2. Длина пластинки рассчитывалась как среднее значение значений, приведенных в литературе или измеренных на гербарных образцах.
Таблица 2 . Морфологические признаки папоротников, использованных в этом исследовании.
Филогенетический сигнал
Чтобы оценить ковариацию эволюционных и морфологических паттернов, мы проверили, проявляют ли выбранные морфологические признаки филогенетический сигнал.Поскольку наше внимание было сосредоточено на функциональном разнообразии, филогенетический сигнал оценивался по всем признакам, а не по отдельным признакам. Поскольку выбранные признаки относятся к смешанным типам данных (категориальным, непрерывным и порядковым), мы сначала вычислили анализ главных координат (PCoA) на основе матрицы несходства Гауэра между всеми парами видов и использовали первую и вторую главные координаты (которые вместе объяснили 51% дисперсии). Мы оценили K Бломберга, используя статистику K (пакет R «Picante»; Бломберг и др., 2003), который измеряет филогенетический сигнал признака, сравнивая наблюдаемый сигнал со случайно развивающимся сигналом филогении. Консерватизм признаков отражается значениями K, близкими к единице или превышающими их, тогда как сходимость признаков отражается значениями, близкими к нулю. Чтобы оценить значимость статистики K, мы создали нулевую модель и случайным образом перемешали метки таксонов 999 раз по концам филогении, вычисляя статистику K для каждой рандомизации и сравнивая ее с дисперсией филогенетически независимых контрастов наблюдаемых данных.
Функциональное разнообразие
Для количественной оценки пространства морфологических признаков (далее «морфопространство»), занимаемого комплексами каждого интервала высот, мы использовали пять индексов функционального разнообразия (дополнительный рисунок S2) с использованием первых пяти осей (80–84% дисперсии) PCoA. вычислено ранее для филогенетических анализов. Мы рассчитали морфообъем с использованием двух дополнительных показателей: функционального богатства (FRic), который использует минимальный выпуклый многоугольник для расчета объема (пакет R «FD»; Villéger et al., 2008), и гиперобъем (Hyp; пакет R «hypervolume»; Blonder et al., 2017), который после процедуры многомерной ядерной оценки плотности очерчивает границы n -мерного объема. FRic чувствителен к выбросам, но может быть вычислен с помощью пяти осей PCoA, а также использовался для оценки упаковки и расширения вдоль разрезов, как указано ниже, тогда как Hyp устойчив к выбросам, но его расчет сложен, поэтому его можно вычислить только с помощью первые две оси PCoA.Затем мы вычислили функциональную равномерность (FEve), которая описывает равномерность распределения в морфопространстве (пакет R «FD»; Villéger et al., 2008). Наконец, мы рассчитали морфоплотность как среднее евклидово расстояние между ближайшими соседями (mNND) и его стандартное отклонение (sdNND).
Поскольку известно, что некоторые из используемых индексов чувствительны к видовому богатству (FRic, NND и SDNN; Mason et al., 2013), мы рассчитали размер стандартизованного эффекта (SES) для каждого показателя с помощью анализа нулевой модели, для которого Названия видов в данных по морфологическим признакам были рандомизированы ( n = 5000 повторов для всех индексов, кроме Hyp с n = 500).Этот подход известен как «независимый обмен» и был предпочтен рандомизации распределения высот видов, поскольку он поддерживает наблюдаемые закономерности богатства и смежности ареала, а также ковариантности признаков и общих фенотипов (Swenson, 2014).
Чтобы понять относительный вклад упаковки и расширения признаков в изменения видового богатства вдоль разрезов, мы количественно оценили расширение ниши и упаковку ниш между соседними высотными полосами. Поскольку видовое богатство представляло собой узор в форме горба, мы определили направление от крайних высот до места, где было достигнуто наибольшее количество видов.Затем мы вычислили процент дополнительных видов между более бедной высотой (E2) и объемом размером E2 внутри более богатой возвышенности (E1), а также количество видов за пределами этого объема. Поскольку сообщества могут отличаться по форме или положению занимаемого объема, мы разработали алгоритм, который, реализуя поиск по сетке, вычисляет процент видов в E1, которые могут содержаться в минимальном выпуклом многоугольнике объема, эквивалентного объему E2. Затем мы приступили к удалению видов до тех пор, пока объем E1 не стал эквивалентным или близким к объему, занимаемому E2.Таким образом, количество видов, удаленных из E1 во время моделирования, представляет собой увеличение богатства из-за расширения признаков, тогда как количество видов, оставшихся в конце моделирования за вычетом единиц в E2, представляет собой дополнительное богатство, приспособленное за счет упаковки признаков (Pigot et al. др., 2016). Упаковка и расширение ниши были рассчитаны для всего набора данных и эпифитных видов отдельно, но не для наземных видов из-за небольшого разброса видового богатства на разрезах.
Проверка гипотез
Мы исследовали взаимосвязь между индексами функционального разнообразия и климатом, чтобы проверить, влияют ли условия окружающей среды на функциональную морфологию этих растений.Далее мы использовали видовое богатство в качестве индикатора конкуренции за нишевое пространство, чтобы оценить биотический компонент фильтров состава сообщества. Мы проанализировали эпифитные и наземные виды отдельно из-за их нишевых и функциональных различий (Kluge, Kessler, 2007). Мы использовали одну микроклиматическую переменную, а именно, покров эпифитных мохообразных на ветвях деревьев, усредненный по высоте, как показатель влажности воздуха (Karger et al., 2012), и четыре макроклиматические переменные из CHELSA V1.2 (Karger et al., 2017, 2018), а именно, среднегодовая температура (MAT), сезонность температуры (TS), годовое количество осадков (AP) и сезонность осадков (PS). Сезонность – это мера изменений в течение года. Чем больше процент, тем больше изменчивость и требуется, чтобы виды приспосабливались к меняющимся условиям окружающей среды.
Чтобы оценить разницу между обработкой всех разрезов вместе или по отдельности, линейный анализ смешанной модели (LMM) был проведен для всех данных, а линейные модели – для каждого разреза отдельно.Поскольку количество точек данных для каждой трансекты было небольшим, мы не смогли использовать все климатические данные, которые мы использовали ниже, поэтому высота над уровнем моря использовалась вместе с видовым богатством по всем индексам.
Для оценки первой гипотезы мы сопоставили климатические данные и богатство видов со всеми индексами, а также каждую SES с LMM, используя разрезы в качестве случайного фактора. Кроме того, мы сравнили климат с богатством видов, чтобы учесть косвенное влияние на индексы. Проведение анализа моделирования структурным уравнением для учета прямых и косвенных влияний на функциональное разнообразие было невозможно из-за ограниченного количества выборочных участков.Чтобы проверить вторую гипотезу, мы применили логистическую регрессию для сопоставления значений упаковки (поскольку они выражены в процентах) и видового богатства, используя разрезы в качестве случайного фактора.
Все модели были рассчитаны с помощью пакета lmer4 в R (Bates et al., 2015). Для выбора модели мы использовали функцию земснаряда в пакете R «MuMIn» (Barton, 2018). Подгонка модели сравнивалась с использованием информационного критерия Акаике (AICc), скорректированного на смещение, и относительная важность переменной во всех моделях была получена путем суммирования ее весов AICc во всех конкурирующих моделях с ΔAICc <2.Величина вариации, объясняемая фиксированными эффектами каждой линейной смешанной модели, была рассчитана с использованием маргинального номера R 2 Накагавы и Шильцета (Nakagawa and Schielzeth, 2013) из пакета MuMIn в R (Barton, 2018). Пакет предоставляет маржинальный R 2 , который соответствует дисперсии, объясняемой фиксированными эффектами, и условный R 2 , который соответствует дисперсии, объясняемой фиксированными и случайными эффектами вместе. Все статистические анализы были реализованы в программе R v.3.4.0 (R Core Team, 2018).
Результаты
Статистика K для филогенетического сигнала всех признаков вместе выявила значения K 0,017 для оси 1 PCoA и 0,011 для оси 2 (оба значения с p <0,001), что указывает на конвергентный паттерн эволюции с низким филогенетическим сигналом.
Связь индексов с видовым богатством и высотой показала значительные различия для отдельных разрезов по сравнению с анализом, проведенным для всех разрезов вместе (Таблица 3), демонстрируя, что тенденции могут быть неверно истолкованы при изучении отдельных разрезов.
Таблица 3 . Регрессионный анализ функционального разнообразия папоротников и ликофитов в зависимости от видового богатства и высоты по четырем тропическим высотным трансектам.
Вообще говоря, функциональное разнообразие, определяемое всеми индексами, наиболее сильно коррелировало с видовым богатством; они стали еще более очевидными при разделении между эпифитами и наземными животными (рис. 2). После того, как влияние видового богатства было устранено путем расчета стандартизированной величины эффекта, объясненные отклонения уменьшились, показывая, что, за некоторыми исключениями, окружающая среда имела незначительное влияние.Когда все виды были проанализированы вместе, наиболее сильная корреляция была обнаружена с переменными, связанными с температурой и микроклиматом. То же самое наблюдалось у наземных животных, тогда как эпифиты сильнее коррелировали с осадками. И наоборот, богатство видов само по себе, когда оно коррелировано с переменными окружающей среды, было наиболее тесно связано с переменными, связанными с водой, а покров мохообразных был наиболее важной переменной для объяснения богатства эпифитов, наземных животных и объединенных данных (Таблица 4; Дополнительная таблица S2 ).
Рисунок 2 . Переменная важность факторов окружающей среды и видового богатства для пяти индексов функционального разнообразия и их стандартизованная величина воздействия (SES). Значения были получены из лучших линейных смешанных моделей (LMM), объясняющих функциональное разнообразие папоротников и ликофитов на четырех тропических высотных разрезах, взвешенных на значениях AICc (ΔAICc <2). LMM использовала набор различных объясняющих переменных, основанных на видовом богатстве, одной микроклиматической переменной (покров мохообразных) и четырех макроклиматических переменных (среднегодовая температура, сезонность температуры, годовые осадки и сезонность осадков), используя разрезы в качестве случайного фактора.Индексы функционального разнообразия характеризуют объем как функциональный объем (FRic) и гиперобъем (Hyp), ровность (FEve) и плотность как среднее (mNND) и стандартное отклонение (sdNND) расстояния до ближайшего соседа. LMM были рассчитаны для всех данных вместе (общие), а затем разделены между эпифитными и наземными папоротниками. Объясненная дисперсия была получена как среднее значение R 2 среди всех ΔAICc <2 моделей.
Таблица 4 . Переменная важность факторов окружающей среды, влияющих на разнообразие видов тропических папоротников и ликофитов.
Morphovolume, количественно оцененный с использованием FRic и Hyp, показал положительную логарифмическую связь с видовым богатством для всех видов, эпифитов и наземных животных (Рисунки 2, 3). В меньшей степени это также было связано с переменными окружающей среды, хотя прямая корреляция с видовым богатством была сильнее. Наблюдалась слабая отрицательная логарифмическая связь между годовым количеством осадков и FRic и положительная линейная корреляция между покровом мохообразных и Hyp, включая все виды.При разделении по формам жизни сезонность осадков была наиболее значимой для эпифитов, тогда как для наземных животных наблюдалась отрицательная связь со средней годовой температурой. Между тем, используя значения SES, только Hyp по-прежнему демонстрирует корреляцию с окружающей средой, особенно с покровом мохообразных (рис. 3).
Рисунок 3 . Индексы функционального разнообразия папоротников и ликофитов в зависимости от видового богатства и переменных окружающей среды для четырех тропических разрезов в Эквадоре (красный), Мексике (зеленый), Папуа-Новой Гвинее (синий) и Уганде (фиолетовый).Линейные смешанные модели (LMM) использовались для сопоставления климатических данных и богатства видов со всеми индексами, а также стандартизированной величины эффекта (SES) каждого с использованием разрезов в качестве случайного фактора. В качестве климатических данных использовались одна микроклиматическая переменная (мохообразный покров на ветвях деревьев; мохообразный покров) и четыре макроклиматические переменные (среднегодовая температура, сезонность температур, годовые осадки и сезонность осадков). Все значения R 2 являются LMM, содержащими только видовое богатство или климатические данные, указанные на графике, и были частью выбранных моделей для анализа важности переменных (наиболее подходящие модели, ΔAICc <2).Напротив, значения, отмеченные ( * ), представляют собой LMM, которые не были выбраны, но все же имеют существенное объяснение дисперсии.
Функциональная равномерность (FEve) была существенно отрицательно связана с видовым богатством логарифмическим способом для всех видов и при разделении по жизненным формам (Рисунок 3; Дополнительный Рисунок S3). Это говорит о формировании кластеров видов с увеличением видового богатства. Несмотря на то, что видовое богатство было более важным, чем другие переменные для всех видов вместе, сезонность температуры стала такой же важной, когда была разделена эпифитами и наземными животными, и все еще была важна для наземных животных при оценке SES FEve (рис. 2).
Морфоплотность, выраженная средним (mNND) и стандартным отклонением (sdNND) расстояния до ближайшего соседа, лучше всего объяснялась видовым богатством (рис. 2). Отрицательная логарифмическая корреляция обоих индексов с видовым богатством (рис. 3) привела к увеличению плотности. В меньшей степени влияли факторы окружающей среды, которые менялись в зависимости от индекса и рассматриваемых данных. Для mNND годовые осадки были наиболее важным предиктором для всех видов вместе взятых и для эпифитов, тогда как наземные обитатели вообще не коррелировали с факторами окружающей среды (рис. 2).SES mNND показала неглубокую зависимость от сезонности осадков для эпифитов (рисунок 2; дополнительный рисунок S3). Напротив, sdNND для всех видов вместе и эпифитов продемонстрировал положительную корреляцию с покровом мохообразных, тогда как наземные животные не показали никакой связи с окружающей средой (Рисунок 2; дополнительные таблицы S3, S4). Несмотря на то, что SES sdNND всех видов вместе коррелировали с окружающей средой, не было никакой корреляции для каких-либо индивидуальных переменных окружающей среды. Более того, когда они разделены формами жизни, никакой корреляции с окружающей средой не наблюдается (рис. 3).
Упаковка положительно логарифмически коррелировала с видовым богатством (рис. 4B: ΔAICc = 28,9 χ ). 2 (1) = 31,6, p <0,001; Дополнительный рисунок S4: ΔAICc = 3,49 χ 2 (1) = 11,4, p <0,001), что показывает увеличение объема при низком уровне видового богатства. Однако он был почти насыщен на более высоких уровнях в результате того, что новые виды в основном упаковывались в уже существующий объем и, таким образом, увеличивалась плотность.
Рисунок 4 . Морфоническая упаковка и разрастание папоротников по четырем высотным градиентам. (A) Процентный вклад упаковки (черный) и расширения (серый) между соседними высотными полосами, показывающий видовое богатство цветными пунктирными линиями. Если нет различий в видовом богатстве между ступенями возвышения, то было невозможно рассчитать индексы, которые отображаются светло-серым цветом как нулевые. (B) Связь процента упаковки с увеличением видового богатства для всех трансект [ΔAICc = 28.9 χ 2 (1) = 31,6, p <0,001]. Трансекты обозначены цветами: Эквадор (красный), Мексика (зеленый), Папуа-Новая Гвинея (синий) и Уганда (фиолетовый). Общий тренд показан черным цветом, а трансекты показаны тем же цветом, что и в (A) .
Рисунок 5 . Схематическое изображение взаимосвязи функционального разнообразия с условиями окружающей среды и видовым богатством тропических эпифитных и наземных папоротников.
Обсуждение
Первой целью нашего исследования было разделить влияние видового богатства и условий окружающей среды на определение функционального разнообразия сообществ папоротников. Предыдущие исследования не смогли провести это различие, потому что они были сосредоточены на отдельных разрезах, где богатство видов и условия окружающей среды тесно взаимосвязаны, что делает невозможным разделение их влияния на функциональное разнообразие (Kluge and Kessler, 2007; Tanaka and Sato, 2015; Carvajal -Hernández et al., 2018). В нашем исследовании, анализируя трансекты по отдельности, мы действительно обнаружили такую же ситуацию (Таблица 3). Однако, когда все трансекты были проанализированы в комбинации (Таблица 3), модели, которые лучше всего объясняли функциональное разнообразие, были теми, которые преимущественно учитывали влияние видового богатства (Рисунок 2). Кроме того, когда влияние видового богатства было исключено с помощью стандартизированного размера эффекта каждого индекса (SES), функциональное разнообразие лишь слабо коррелировало с факторами окружающей среды.Также важно отметить, что признаки, использованные в этом исследовании, показали низкий филогенетический сигнал, так что взаимосвязь между функциональным разнообразием и видовым богатством определяется не напрямую филогенетическими паттернами, а конвергентными эволюционными паттернами.
Что касается второй цели нашего исследования, мы обнаружили, что функциональный морфообъем почти достиг насыщения с увеличением видового богатства. Полное насыщение будет достигнуто, когда будут реализованы все крайние возможные комбинации, так что новые виды увеличат избыточность или еще больше разделят нишевое пространство, которое уже занято (Mayfield et al., 2010; Cadotte et al., 2011). Мы обнаружили, что при низком уровне видового богатства, в нашем случае на крайних точках каждой трансекты, дополнительные виды в основном имеют тенденцию к увеличению объема. Напротив, с увеличением видового богатства по направлению к средним высотам, добавление видов было в основном за счет увеличения упаковки (рис. 4). Свенсон и Вайзер (2014) представляют аналогичные результаты для сообществ деревьев в восточной части Северной Америки, где как объем, так и степень упаковки увеличивались с увеличением видового богатства.В своем исследовании андских птиц Pigot et al. (2016) также заметили, что высокое видовое богатство сильнее связано с более плотным заполнением морфопространства признака, чем с увеличением объема ниши.
При более внимательном рассмотрении показателей упаковки ниш мы обнаружили, что среднее и стандартное отклонения расстояний до ближайших соседей, а также равномерность уменьшаются с увеличением видового богатства. Это указывает на то, что виды упакованы в одних частях объема, тогда как другие заняты редко, что может быть результатом ковариаций между изучаемыми признаками, так что не все комбинации признаков возможны.Более того, увеличение однородности с богатством видов часто интерпретируется как ограничение фенотипического сходства (Kraft et al., 2008). Поскольку ровность показала снижение, наши результаты не подтверждают этого ограничения, как это было ранее обнаружено для андских птиц (Pigot et al., 2016).
Хотя мы обнаружили, что видовое богатство было самым сильным определяющим фактором морфологического разнообразия в наших исследуемых сообществах, мы также обнаружили, что факторы окружающей среды играли второстепенную роль, особенно когда мы проводили различие между формами жизни (Рисунок 2; SES индексов).Эпифиты чаще подвергаются частым и даже продолжительным периодам засухи, чем наземные животные, живущие в той же среде обитания (Watkins et al., 2007). Чем выше в кроне деревьев, тем сильнее интенсивность света и меняется влажность воздуха, при этом мало почвы служит буфером. Даже небольшие изменения условий окружающей среды могут оказать значительное влияние на растения, обитающие в пологе (Petter et al., 2016). Наши результаты согласуются с этим и показывают, что на разнообразие признаков у эпифитов в основном влияет сезонность, в частности сезонность осадков.Для наземных папоротников наиболее существенное влияние оказали температурные переменные. Поскольку в тропиках высокая влажность, а почва является буфером для микроклиматических изменений, наиболее существенные колебания, которым подвергаются наземные виды, будут вызваны изменениями температуры. Однако картина меняется, если вместо изучения влияния на разнообразие функциональных признаков принимается во внимание влияние окружающей среды на видовое богатство. Наши результаты показали, что, хотя видовое богатство показало сходные закономерности с функциональным разнообразием, наиболее важным фактором, независимо от формы жизни, была микроклиматическая переменная, а именно эпифитный покров мохообразных, который мы использовали в качестве показателя влажности воздуха.Фактически, разнообразие папоротников в первую очередь обусловлено водно-энергетическим балансом, при этом наличие воды является наиболее ограничивающим фактором (Kreft et al., 2010; Qian et al., 2012; Khine et al., 2019; Weigand et al., 2020). Вместе это раскрывает косвенное влияние на окружающую среду через богатство видов, а не разнообразие признаков (рис. 5).
Как и любое исследование подобного рода, у нашего есть методологические ограничения. Во-первых, мы были ограничены морфологическими признаками, которые можно легко собрать для большого числа видов (всего 800 в нашем исследовании), а не физиологическими и анатомическими признаками, такими как масса листа на площадь (LMA), что типично для исследований функциональных возможностей растений. черты характера (e.г., Bruelheide et al., 2018). Нет сомнений в том, что такие признаки, как размер растения или пространственное распределение системы корневищ, являются адаптивными и играют роль в приобретении ресурсов и межвидовой конкуренции. Тем не менее, наше исследование, возможно, дало бы другие результаты, если бы мы включили другой набор черт. Тем не менее, мы считаем это маловероятным, поскольку исследование бразильских папоротников с использованием признаков экономического спектра листьев также обнаружило сильный сигнал конвергенции признаков по градиентам окружающей среды, что согласуется с нашими результатами (Costa et al., неопубликованные данные). Во-вторых, большинство наших черт были оценены как категориальные или порядковые, отчасти потому, что некоторые черты были бинарными по своей природе, а отчасти потому, что некоторые из них обладают высокой внутривидовой изменчивостью, которая лучше всего отражается в категориях. Этот тип данных может привести к потере разрешения, что может ограничить наши выводы. Однако мы сомневаемся, что на наш главный результат – преобладание видового богатства – повлияла бы более точная оценка морфологического разнообразия. В более общем плане игнорирование внутривидовой изменчивости признаков ограничивает нашу способность различать, были ли изменения в степени упаковки следствием разной ширины ниши признаков (т.е., более тонкая специализация), или совпадение ниш по разным признакам (MacArthur, 1965). Внутривидовая изменчивость признаков актуальна для растительных сообществ в локальных масштабах и вдоль коротких экологических градиентов (Carmona et al., 2015; Siefert et al., 2015), а более тонкая специализация может также лежать в основе градиентов высотного разнообразия (Hulshof et al., 2013). Действительно, предыдущие исследования реализованных экологических ниш среди папоротников показали, что они уменьшаются с увеличением видового богатства (Karger et al., 2015). Наконец, выбранные нами области исследования ограничены тропической средой.Тем не менее, основываясь на нашем понимании глобальных закономерностей разнообразия папоротников (Weigand et al., 2020) и того факта, что данные графика разрезов можно использовать для экстраполяции структур богатства на большие географические регионы (Khine et al., 2019), мы уверены что наши трансекты репрезентативны и что наши результаты широко применимы к ассоциациям тропических континентальных папоротников. Применимы ли они также к скоплениям папоротников умеренного пояса и океанических островных зон, еще предстоит изучить.
Взятые вместе и с учетом вышеупомянутых ограничений, наши результаты показывают, что высокое видовое богатство прогрессивно способствует упаковке морфологического пространства ниши, в то же время увеличивая функциональное разнообразие.Более того, при заданном уровне видового богатства окружающая среда не сильно изменяла морфологическую структуру ниши, хотя в нашем случае это часто происходило в различных климатических условиях на трансектах, принадлежащих к разным биогеографическим регионам с сильно отличающимся региональным разнообразием папоротников (Kreft et al. ., 2010; Weigand et al., 2020). Поскольку на видовое богатство само по себе сильно влияют условия окружающей среды (Wright, 1983; Hawkins et al., 2003; Kessler et al., 2011), существует косвенное влияние окружающей среды на разнообразие признаков.Тем не менее, основной движущей силой стратегий получения ресурсов папоротников, по-видимому, является количество видов.
Заявление о доступности данных
Необработанные данные, подтверждающие выводы этой статьи, будут предоставлены авторами без излишних оговорок.
Авторские взносы
MK, ML и DQ разработали идеи и разработали методологию. Данные собирали AH-R, CC-H, DA-M, DK, DQ, JK, MK, ML, MS, LS и SN. DA-M, DK и SN проанализировали данные.DA-M и MK руководили написанием рукописи. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Это исследование проводилось при поддержке проектов MK Швейцарского национального научного фонда (310030L 146906), DK (148691 и 20BD21_184131 / 1) и проекта ML (LE1826 / 4 и LE1826 / 5) и DQ (QU153) Немецкого исследовательского фонда. / 7).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы благодарим Rose Badaza и Voytech Novotny за поддержку полевых работ, власти стран исследования за предоставление разрешений на исследования и Rafael O. Wüest-Karpati за поддержку в статистическом анализе.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2020.615723/full#supplementary-material
Список литературы
Бейтс, Д., Махлер, М., Болкер, Б.М., и Уокер, С.С. (2015). Подгонка линейных моделей смешанных эффектов с использованием lme4. J. Stat. Софтв. 67, 1–48. DOI: 10.18637 / jss.v067.i01
CrossRef Полный текст | Google Scholar
Бломберг, С. П., Гарланд, Т., и Айвз, А. Р. (2003). Проверка на филогенетический сигнал в сравнительных данных: поведенческие черты более лабильны. Evolution 57, 717–745. DOI: 10.1111 / j.0014-3820.2003.tb00285.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Блондинка, Б., Морроу, К. Б., Мейтнер, Б., Харрис, Д. Дж., Ламанна, К., Виолле, К., и др. (2017). Новые подходы к разграничению n-мерных гиперобъемов. Methods Ecol. Evol. 9, 305–319. DOI: 10.1111 / 2041-210X.12865
CrossRef Полный текст | Google Scholar
Брюлхайде, Х., Денглер, Дж., Пуршке, О., Ленуар, Дж., Хименес-Альфаро, Б., Хеннекенс, С. М., и др. (2018). Глобальные взаимоотношения растительных сообществ между признаком и окружающей средой. Нат. Ecol. Evol. 2, 1906–1917. DOI: 10.1038 / s41559-018-0699-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кадотт, М. В., Карскадден, К., Мирочник, Н. (2011). Помимо видов: функциональное разнообразие и поддержание экологических процессов и услуг. J. Appl. Ecol. 48, 1079–1087. DOI: 10.1111 / j.1365-2664.2011.02048.x
CrossRef Полный текст | Google Scholar
Кармона, К. П., Рота, К., Азкарат, Ф. М., и Пеко, Б. (2015). Больше за меньшие деньги: стратегии выборки функциональных характеристик растений в зависимости от местных градиентов окружающей среды. Funct. Ecol. 29, 579–588. DOI: 10.1111 / 1365-2435.12366
CrossRef Полный текст | Google Scholar
Карвахаль-Эрнандес, К. И., Гомес-Диас, Дж. А., Кесслер, М., и Кремер, Т. (2018). Влияние высот и нарушения среды обитания на функциональное разнообразие папоротников и ликофитов. Plant Ecol. Дайверы. 11, 335–347. DOI: 10.1080 / 17550874.2018.1484526
CrossRef Полный текст | Google Scholar
Колвелл, Р. К. (2013). EstimateS: статистическая оценка видового богатства и общих видов по выборкам.Версия 9 и более ранние. Руководство пользователя и приложение. Доступно по адресу: http://purl.oclc.org/estimates (по состоянию на 20 июня 2018 г.).
Google Scholar
Диас, С., Кабидо, М. (2001). Vive la différence: функциональное разнообразие растений имеет значение для экосистемных процессов. Trends Ecol. Evol. 16, 646–655. DOI: 10.1016 / S0169-5347 (01) 02283-2
CrossRef Полный текст | Google Scholar
Гарсон-Лопес, К. X., Бальестерос-Мехиа, Л., Ордоньес, А., Больман, С. А., Ольф, Х., и Янсен, П. А. (2015). Косвенное взаимодействие между видами тропических деревьев через общих хищников семян грызунов: новый механизм сосуществования видов деревьев. Ecol. Lett. 18, 752–760. DOI: 10.1111 / ele.12452
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Грайм, Дж. П. (2006). Конвергенция и расхождение признаков в сообществах травянистых растений: механизмы и последствия. J. Veg. Sci. 17, 255–260. DOI: 10.1111 / j.1654-1103.2006.tb02444.x
CrossRef Полный текст | Google Scholar
Хокинс, Б. А., Филд, Р., Корнелл, Х. В., Карри, Д. Дж., Геган, Дж. -Ф., Кауфман, Д. М. и др. (2003). Энергия, вода и широкомасштабные географические закономерности видового богатства. Экология 84, 3105–3117. DOI: 10.1890 / 03-8006
CrossRef Полный текст | Google Scholar
Херцог, С. К., Кесслер, М., и Кэхилл, Т. М. (2002). Оценка видового богатства сообществ тропических птиц по данным экспресс-оценки. Auk 119, 749–769. DOI: 10.1093 / auk / 119.3.749
CrossRef Полный текст | Google Scholar
Холттум Р. Э. (1991). Flora Malesiana, серия II, Pteridophyta, т. 2, часть 1, группа Tectaria. Рейксербарий / Hortus Botanicus, Лейден.
Google Scholar
Hulshof, C. M., Violle, C., Spasojevic, M. J., McGill, B., Damschen, E., Harrison, S., et al. (2013). Внутривидовые и межвидовые вариации в определенной площади листа показывают важность абиотических и биотических факторов видового разнообразия на высоте и широте. J. Veg. Sci. 24, 921–931. DOI: 10.1111 / jvs.12041
CrossRef Полный текст | Google Scholar
Хатчинсон, Г. (1957). Популяционные исследования – экология и демография животных – заключительные замечания. Cold Spring Harb. Symp. Quant. Биол. 22, 415–427. DOI: 10.1101 / SQB.1957.022.01.039
CrossRef Полный текст | Google Scholar
Каргер, Д. Н., Конрад, О., Бёнер, Дж., Каволь, Т., Крефт, Х., Сориа-Ауза, Р. В. и др. (2017). Климатология с высоким разрешением для участков земной поверхности. Sci. Данные 4: 170122. DOI: 10.1038 / sdata.2017.122
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каргер, Д. Н., Конрад, О., Бёнер, Дж., Каволь, Т., Крефт, Х., Сориа-Ауза, Р. В. и др. (2018). Данные из: Climatologies в высоком разрешении для участков земной поверхности. Набор данных Dryad . DOI: 10.5061 / dryad.kd1d4
CrossRef Полный текст | Google Scholar
Каргер, Д. Н., Клюге, Дж., Стефан, А., Салазар, Л., Хомейер, Дж., Lehnert, M., et al. (2012). Мохообразный покров деревьев как показатель влажности воздуха в тропиках. Ecol. Инд. 20, 277–281. DOI: 10.1016 / j.ecolind.2012.02.026
CrossRef Полный текст | Google Scholar
Каргер Д. Н., Туомисто Х., Аморосо В. Б., Дарнаеди Д., Хидаят А., Абрахамчик С. и др. (2015). Важность размера пула видов для состава сообщества. Экография 38, 1243–1253. DOI: 10.1111 / ecog.01322
CrossRef Полный текст | Google Scholar
Каргер, Д.N., Weigelt, P., Amoroso, V. B., Darnaedi, D., Hidayat, A., Kreft, H., et al. (2014). Биогеография острова в масштабах от регионального до местного: свидетельство пространственно масштабированной эхо-модели разнообразия папоротников на архипелаге Юго-Восточной Азии. J. Biogeogr. 41, 250–260. DOI: 10.1111 / jbi.12209
CrossRef Полный текст | Google Scholar
Кесслер М. и Бах К. (1999). Использование индикаторных семейств для классификации растительности в неотропических лесах с богатыми видами. Phytocoenologia 29, 485–502.
Google Scholar
Кесслер М., Клюге Дж., Хемп А. и Олемюллер Р. (2011). Глобальный сравнительный анализ образцов высокогорного видового богатства папоротников. Glob. Ecol. Биогеогр. 20, 868–880. DOI: 10.1111 / j.1466-8238.2011.00653.x
CrossRef Полный текст | Google Scholar
Хин, П. К., Клюге, Дж., Кесслер, М., Миэ, Г., и Каргер, Д. Н. (2019). Не зависящая от широты, общеконтинентальная стабильность взаимосвязей между климатом и богатством азиатских папоротников и ликофитов. J. Biogeogr. 46, 981–991. DOI: 10.1111 / jbi.13558
CrossRef Полный текст | Google Scholar
Клюге, Дж., И Кесслер, М. (2007). Морфологическая характеристика скоплений папоротников по наклонной высоте: закономерности и причины. Ecotropica 13, 27–43.
Google Scholar
Крафт, Н. Дж. Б., Валенсия, Р., и Акерли, Д. Д. (2008). Функциональные особенности и сборка сообществ деревьев на основе ниши в лесах Амазонки. Наука 322, 580–582.DOI: 10.1126 / science.1160662
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крефт, Х., Джетц, В., Мутке, Дж., И Бартлотт, В. (2010). Противопоставление экологического и регионального воздействия на глобальное разнообразие птеридофитов и семенных растений. Экография 33, 408–419. DOI: 10.1111 / j.1600-0587.2010.06434.x
CrossRef Полный текст | Google Scholar
Ламанна, К., Блондер, Б., Виолле, К., Крафт, Н. Дж. Б., Сандель, Б., Шимова, И., и др. (2014).Пространство функциональных признаков и градиент широтного разнообразия. Proc. Natl. Акад. Sci. 111, 13745–13750. DOI: 10.1073 / pnas.1317722111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
MacArthur, R., and Levins, R. (1967). Предельное сходство, конвергенция и расхождение сосуществующих видов. Am. Nat. 101, 377–385. DOI: 10.1086 / 282505
CrossRef Полный текст | Google Scholar
Мейсон, Н. В. Х., де Белло, Ф., Муийо, Д., Павуан, С., Дрей, С. (2013). Руководство по использованию индексов функционального разнообразия для выявления изменений в процессах сборки по экологическим градиентам. J. Veg. Sci. 24, 794–806. DOI: 10.1111 / jvs.12013
CrossRef Полный текст | Google Scholar
Мэйфилд, М., Бонсер, С. П., Морган, Дж. У., Обин, И., Макнамара, С., и Веск, П. А. (2010). Что богатство видов говорит нам о разнообразии функциональных признаков? Прогнозы и доказательства реакции видов и разнообразия функциональных признаков на изменения в землепользовании. Glob. Ecol. Биогеогр. 19, 423–431. DOI: 10.1111 / j.1466-8238.2010.00532.x
CrossRef Полный текст | Google Scholar
Мишале Р. и Пуньер Ф. И. (2016). Содействие в сообществах: основные механизмы, последствия для сообществ и экосистем. Funct. Ecol. 30, 3–9. DOI: 10.1111 / 1365-2435.12602
CrossRef Полный текст | Google Scholar
Микель Дж. И Смит А. Р. (2004). Птеридофиты Мексики . Нью-Йорк: Пресса ботанического сада Нью-Йорка.
Google Scholar
Моше, М. А., Виллегер, С., Мейсон, Н. В. Х. и Муийо, Д. (2010). Меры функционального разнообразия: обзор их избыточности и их способности различать правила собрания сообщества. Funct. Ecol. 24, 867–876. DOI: 10.1111 / j.1365-2435.2010.01695.x
CrossRef Полный текст | Google Scholar
Накагава С., Шильцет Х. (2013). Общий и простой метод получения R2 из обобщенных линейных моделей смешанных эффектов. Methods Ecol. Evol. 4, 133–142. DOI: 10.1111 / j.2041-210x.2012.00261.x
CrossRef Полный текст | Google Scholar
Нитта, Дж. Х., Уоткинс, Дж. Э. и Дэвис, К. С. (2020). Жизнь в пологе: оценка особенностей сообщества выявляет значительное функциональное разнообразие эпифитов папоротников. New Phytol. 227, 1885–1899. DOI: 10.1111 / Nph.16607
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Петтер, Г., Вагнер, К., Ванек, В., Дельгадо, Э.J. S., Zotz, G., Cabral, J. S., et al. (2016). Функциональные признаки листьев сосудистых эпифитов: вертикальные тенденции в пределах леса, внутри- и межвидовая изменчивость признаков и таксономические сигналы. Funct. Ecol. 30, 188–198. DOI: 10.1111 / 1365-2435.12490
CrossRef Полный текст | Google Scholar
Пигот А. Л., Трисос К. Х. и Тобиас Дж. А. (2016). Функциональные черты показывают расширение и упаковку пространства экологической ниши, лежащую в основе градиента высотного разнообразия воробьиных птиц. Proc. R. Soc. B Biol. Sci. 283: 20152013. DOI: 10.1098 / rspb.2015.2013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Цянь, Х., Ван, С., Ли, Ю., Сяо, М., и Ван, X. (2012). Разоблачение относительного воздействия энергии окружающей среды, доступности воды и водно-энергетического баланса на богатство видов птеридофитов в ландшафтном масштабе в Китае. Plant Ecol. 213, 749–756. DOI: 10.1007 / s11258-012-0038-0
CrossRef Полный текст | Google Scholar
R Основная команда (2018). Язык и среда для статистических вычислений . Вена, Австрия: Фонд статистических вычислений.
Google Scholar
Риклефс, Р. Э., и Кокс, Г. У. (1977). Морфологическое сходство и экологическое совпадение у воробьиных птиц на острове Сент-Китс, Британская Вест-Индия. Oikos 29, 60–66. DOI: 10.2307 / 3543293
CrossRef Полный текст | Google Scholar
Розенфельд, Дж. С. (2002). Функциональная избыточность в экологии и сохранении. Oikos 98, 156–162. DOI: 10.1034 / j.1600-0706.2002.980116.x
CrossRef Полный текст | Google Scholar
Siefert, A., Violle, C., Chalmandrier, L., Albert, C.H., Taudiere, A., Fajardo, A., et al. (2015). Глобальный метаанализ относительной степени вариации внутривидовых признаков в растительных сообществах. Ecol. Lett. 18, 1406–1419. DOI: 10.1111 / ele.12508
CrossRef Полный текст | Google Scholar
Штольце, Р. Г., и Пачеко, Л. (1994).Флора Эквадора. Polypodiaceae, Dryopteridoideae, Physematieae (том 49–53). Ботанический институт Гетеборгского университета.
Google Scholar
Свенсон, Н. (2014). Функциональная и филогенетическая экология в R . Нью-Йорк: Springer-Verlag.
Google Scholar
Свенсон, Н. Г., и Вейзер, М. Д. (2014). Об упаковке и заполнении функционального пространства в восточно-североамериканских сообществах деревьев. Экография 37, 1056–1062. DOI: 10.1111 / ecog.00763
CrossRef Полный текст | Google Scholar
Танака Т., Сато Т. (2015). Таксономическое, филогенетическое и функциональное разнообразие папоротников и ликофитов вдоль градиента высоты зависит от таксономических шкал. Plant Ecol. 216, 1597–1609. DOI: 10.1007 / s11258-015-0543-z
CrossRef Полный текст | Google Scholar
Verdcourt, B., Roux, P., and Shaffer-Fehre, M. (1999/2008). Флора тропической Восточной Африки . Королевский ботанический сад, Кью.
Google Scholar
Виллегер С., Мейсон Н. В. Х. и Муйо Д. (2008). Новые многомерные индексы функционального разнообразия для многогранной основы функциональной экологии. Экология 89, 2290–2301. DOI: 10.1890 / 07-1206.1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Уоткинс, Дж. Э., Рундель, П. У. и Карделус, К. Л. (2007). Влияние жизненной формы на отношения углерода и азота в папоротниках тропических лесов. Oecologia 153, 225–232.DOI: 10.1007 / s00442-007-0723-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Weigand, A., Abrahamczyk, S., Aubin, I., Bita-Nicolae, C., Bruelheide, H., Carvajal-Hernández, C. I., et al. (2020). Объяснение глобального богатства папоротников и ликофитов: как региональные и местные факторы влияют на богатство участков. J. Biogeogr. 47, 59–71. DOI: 10.1111 / jbi.13782
CrossRef Полный текст | Google Scholar
Вейхер Э. и Кедди П. А. (1995). Правила сборки, нулевые модели и дисперсия признаков: новые вопросы из старых шаблонов. Oikos 74, 159–164. DOI: 10.2307 / 3545686
CrossRef Полный текст | Google Scholar
Вольф П. Г., Шнайдер Х. и Ранкер Т. А. (2001). Географическое распространение гомоспористых папоротников: скрывает ли расселение свидетельство викариантности? J. Biogeogr. 28, 263–270. DOI: 10.1046 / j.1365-2699.2001.00531.x
CrossRef Полный текст | Google Scholar
сортов папоротниковых растений | Home Guides
Папоротник подходит для любого места в вашем саду: солнце, глубокая тень, влажная или сухая почва, альпинарии и даже на стволах деревьев.Они растут во всех размерах. Древесные папоротники могут достигать 20 футов, в то время как некоторые разновидности папоротников девичьей шерсти достаточно крошечные, чтобы поместиться в террариумах. Папоротники могут быть как вечнозелеными, так и листопадными. Выявлено более 12 000 видов папоротников.
Древовидные папоротники
Древесные папоротники, многие виды которых существуют в пределах родов Dicksonia и Cyathea, обычно имеют один прямой ствол и зонтик из ветвей, выходящих из верхушки. Некоторые виды морозоустойчивы, но большинство из них следует либо завернуть, либо принести внутрь, если ожидаются отрицательные температуры.Они могут расти в земле в средиземноморском климате и в горшках где угодно.
Папоротники, любящие тень
Большинство папоротников предпочитают влажную, тенистую среду. Один культивируемый папоротник, который растет в тени, – это рождественский папоротник (Polystichum acrostichoides), густой сорт, получивший свое название за то, что он остается зеленым в праздничные дни. Рождественский папоротник достигает от 1,5 до 2 футов в высоту и является морозостойким в зонах 3–9 Министерства сельского хозяйства США. Леди папоротник (Athyrium filix-femina), имеющий кружевные листья и распространенный по всей территории Соединенных Штатов, вырастает от 2 до 3. футов высотой во влажной тени и выносливы в зонах USDA с 4 по 9.Еще один любитель оттенков – папоротник меченосный (P. munitum), у которого прямостоячие листья до 6 футов высотой. Это обычное явление в западных штатах и морозостойкое в зонах USDA с 8 по 11.
Необычные папоротники
По крайней мере, у двух разновидностей папоротников нет обычных многосегментных листьев, которые обычно ассоциируются с папоротниками. Папоротники оленьего рога (Platycerium spp.) Можно сразу узнать по их драпированным раздвоенным ветвям, имеющим форму рогов, и круглым базальным структурам, которые прикрепляются к деревьям, камням и другим поверхностям.Домашние производители часто помещают их в проволочные корзины, чтобы их можно было перемещать в помещении при понижении температуры. Растения оленьего рога могут вырасти до нескольких футов в диаметре и до нескольких сотен фунтов в весе. Поскольку они эпифиты, или воздушные растения, им не нужна почва; вместо этого они получают питание от влажности воздуха. Оленьорог процветает в зоне 10 USDA. Папоротник птичьего гнезда (Asplenium nidus) вырастает до 4 футов в высоту и ширину в тени или полутени. Его длинные листья исходят из центрального пучка, что дало папоротнику свое название.Папоротники «птичьи гнезда» выносливы в зонах USDA с 9 по 11.
Папоротники другого цвета
Не все папоротники зеленые. Бразильский древовидный папоротник (Blechnum brasiliense) в молодом возрасте дает листья с красными кончиками. Это один из небольших древовидных папоротников, вырастающий от 1 до 2 футов в высоту, и он вынослив в зоне 10 USDA. Пестролистный папоротник (Pteris ensiformis “Evergemiensis”) имеет тонкие зеленые листья с белыми центрами, которые составляют около 1 фута в высоту. Он морозостойкий в зонах USDA с 9 по 12. Вирджинский голубой папоротник (Polypodium pseudo-aureum) имеет голубоватые листья, вырастает от 1 до 2 футов в высоту и хорошо растет в зонах USDA с 8 по 10.
Ссылки
Ресурсы
Писатель Биография
Патрисия Роквуд была профессиональным редактором и писателем более 25 лет. Она заядлый садовник с сертифицированной средой обитания на заднем дворе Флориды. Роквуд занимается йогой более 40 лет и большую часть этого времени преподает. Она также профессиональный художник-мозаичист.
Девятнадцать видов папоротников, названных в честь Леди Гаги
Леди Гага, мегазвезда поп-музыки, удостоена звания нового рода папоротников, произрастающих в Центральной и Южной Америке, Мексике, Аризоне и Техасе.Род – это группа близкородственных видов; в этом случае 19 видов папоротников будут носить название Гага.
На одном из этапов своей жизни новый род Гага имеет несколько плавное определение пола и имеет поразительное сходство с одним из знаменитых костюмов Гаги. Члены нового рода также несут отчетливую последовательность ДНК, пишущую GAGA.
Два вида рода Gaga являются новыми для науки: Gaga germanotta из Коста-Рики названа в честь семьи художника, рожденного Стефани Джерманотта.А недавно обнаруженный мексиканский вид получил название Gaga monstraparva (буквально монстр-маленький) в честь поклонников Гаги, которых она называет «маленькими монстрами».
«Мы хотели назвать этот род по имени Леди Гага, потому что она яростно защищала равенство и индивидуальное самовыражение», – сказала руководитель исследования Кэтлин Прайер, профессор биологии Университета Дьюка и директор Герцогского гербария. «И когда мы начали рассматривать это, сами папоротники дали нам больше причин, почему это был хороший выбор».
Например, в своем выступлении на церемонии вручения премии Грэмми 2010 Леди Гага была одета в костюм Армани Прайва в форме сердца с гигантскими плечами, которые для натренированного глаза Прайер выглядели точно так же, как бисексуальная репродуктивная стадия папоротников, называемая гаметофитом.Это был даже правильный оттенок светло-зеленого. То, как папоротник распускает свои новые листья в виде сжатого маленького шарика, также напоминает Прайер о том, как Гага приветствует своих поклонников «лапками вверх».
Решающий аргумент пришел, когда аспирантка Фэй-Вэй Ли просканировала ДНК папоротников, считающихся отнесенными к новому роду. Он обнаружил, что GAGA записана в парах оснований ДНК как признак, который отличает эту группу папоротников от всех остальных.
Виды знаменитостей изобилуют наукой. В Калифорнии есть лишайник, названный в честь президента Барака Обамы, и мясоедное растение в джунглях, названное в честь актрисы Хелен Миррен.В январе австралийская конная муха, которую ее первооткрыватель назвал “добычей”, была названа в честь певицы Бейонсе.
Леди Гага и гаметофит папоротника.
Но это всего лишь отдельные виды. Это целый род, который на данный момент включает 19 видов папоротников.
За исключением двух новых видов, germanotta и monstraparva, остальные папоротники Gaga – это виды, которые Прайер и ее соавторы реклассифицируют. Ранее они были отнесены к роду Cheilanthes на основании их внешнего вида.Но тщательный анализ ДНК, проведенный Ли с использованием более 80 музейных образцов и недавно собранных растений, показал, что они отличаются друг от друга и заслуживают своего собственного рода.
Новые инструменты генетического анализа реорганизуют генеалогическое древо папоротников, сказала Прайер, которая в настоящее время является президентом Американского общества папоротников и президентом Американского общества систематиков растений, ученых, которые называют и классифицируют виды растений.
Как и большинство папоротников, группа Гага «гомоспористая». Они производят крошечные сферические споры, которые падают на землю и прорастают в сердцевидные растения, называемые гаметофитами.Эти независимые маленькие организмы могут быть женскими, мужскими или даже бисексуальными, в зависимости от условий роста и других видов гаметофитов. При подходящих условиях они обмениваются спермой между гаметофитами, но при необходимости они также могут самооплодотворяться, чтобы произвести новый папоротник.
«Биология этих папоротников чрезвычайно неясна и размыта из-за полового скрещивания между видами», – сказала Прайер. “У них большое количество хромосом и асексуальность, что может привести к потомству, которое генетически идентично родительскому растению.«
Прайер открыто признает, что она и ее лаборатория – большие фанаты Гаги. «Мы часто слушаем ее музыку, пока проводим свои исследования. Мы думаем, что ее второй альбом« Born this Way »чрезвычайно расширяет возможности, особенно для обездоленных людей и сообществ, таких как ЛГБТ, этнические группы, женщины – и ученых, изучающих странные вещи. папоротники! ” – сказала Прайер.
«Какая замечательная, неожиданная, совершенная дань уважения названию рода папоротников для Леди Гаги», – сказала член факультета Герцога Кэти Н. Дэвидсон, которая участвовала в Инициативе Фонда Макартура по цифровым медиа и обучению, которая помогла Леди Гаге создать Рожденного This Way Foundation, национальная инициатива по борьбе с издевательствами.«Поощрять своих поклонников и детей повсюду быть смелыми, смелыми, уникальными, творческими и умными – вот что делает Леди Гага», – говорит Дэвидсон. «Редко, когда такая молодая знаменитость отдает так много обществу».
Исследование частично финансировалось Национальным научным фондом, грант DEB-0717398.
ЦИТАТА: «Гага, новый род папоротников, отделенный от Cheilanthes (Pteridaceae)», Фэй-Вей Ли, Кэтлин М. Прайер и Майкл Д. Виндхэм. Систематическая ботаника, октябрь-декабрь 2012 г.
«Странный» папоротник демонстрирует силу межвидового пола: NPR
Ботаники говорят, что это растение – папоротник, эквивалентный ребенку любви человека-лемура. Гарри Роскам скрыть подпись
переключить подпись Гарри РоскамБотаники говорят, что это растение – папоротник, похожий на дитя любви человека и лемура.
Гарри РоскамКажется, любовь между двумя папоротниками не знает границ.Анализ ДНК гибридного папоротника показывает, что его родители – это два разных вида, разделенных почти 60 миллионами лет эволюции.
«Расхождение в 60 миллионов лет приблизительно эквивалентно спариванию человека с лемуром», – говорит Карл Ротфельс, исследователь папоротников из Университета Британской Колумбии, возглавлявший исследование. По его словам, гибрид – это рекорд.
Необычный папоротник, известный как Cystocarpium roskamianum, , не является редкостью. Его можно найти раскинувшимся во французских Пиренеях.По словам Ротфельса, его даже можно купить в некоторых европейских садовых центрах.
Но исследователи папоротников всегда думали, что это выглядит довольно странно.
«Я имею в виду, что с одной стороны это похоже на папоротник», – признает Ротфельс.
Но он говорит, что этот сорт папоротника, похоже, произошел от двух родителей, которых вы не ожидали бы стать парой.Один живет на скалистых обнажениях. Другой находится на подстилках леса. Это два разных вида из разных мест, но каким-то образом они собрались вместе, чтобы создать этот гибрид.
«Это почти точно между двумя родителями», – говорит он.
Анализ ДНК группы, подтверждающий нечетное сцепление, опубликован в мартовском выпуске журнала The American Naturalist .
«Папоротники уникальны среди растений по многим причинам, и это исследование добавляет еще одно потенциальное отличие к списку», – говорит Эмили Сесса, исследователь из Университета Флориды в Гейнсвилле.По-видимому, у папоротников не так легко возникают барьеры, которые мешают им скрещиваться.
Эта гибкость может помочь объяснить, почему на планете всего 10 000 видов папоротников по сравнению с примерно 250 000 видов цветковых растений, добавляет Роббин Моран, куратор папоротников в ботаническом саду Нью-Йорка.
Ротфельс говорит, что гибридный папоротник бесплоден, хотя он может воспроизводиться «вегетативно», рассылая побеги по земле.
«Кажется, вполне устраивает», – говорит он.
Красивых и необычных сортов папоротников для сада
Когда я впервые влюбился в тогда еще редкий осенний папоротник ( Dryopteris erythrosora , USDA Hardiness Zones 5–9) еще в конце 1960-х годов, папоротники были чужими в саду, и лишь горстка местных жителей была выставлена на продажу. мало признания их ценности дизайнерами или даже преданными садоводами. Все изменилось, и теперь есть выбор для любых целей, от миниатюрных образцов альпинария до почвенных покровов и высоких стражей, которые создают координационные центры.Некоторые из них вечнозеленые, а некоторые лиственные, некоторые принимают солнце, но большинство предпочитают тень, а некоторые переносят засуху, а другие предпочитают влажные и заболоченные места. Листья бывают бесконечных вариантов текстуры и формы – кружевные, изящные, смелые, мягкие, прямые или рыхлые, а также различных оттенков цвета, включая розовые оттенки. Следующие папоротники – одни из моих любимых. Все они вечнозеленые, поэтому будут очаровывать вас круглый год. Обратите внимание, что размеры меняются в зависимости от зоны, почвы, воды и воздействия.
Цепной папоротник, украшенный драгоценностями, уникальный
Имя: Woodwardia unigemmata
Зоны: с 8 по 10
Размер: Стебли и листья длиной до 6 футов, каскадные
Условия: полутень; влажная почва
Цепной папоротник, украшенный драгоценными камнями, несомненно, является драгоценным камнем с эффектными изогнутыми ветвями, которые появляются в ярких оттенках красного и легко ниспадают каскадом по стене или склону. Каждая вайя вырастает до 6 футов в длину и заканчивается луковицей, готовой к укоренению, которую вайя пытается опустить в приемлемую почву.Эти луковицы можно собирать для размножения в конце сезона, когда они пухлые. Поместите их на влажную, но не мокрую почву в небольшой горшок, накройте горшок прозрачным пластиковым стаканчиком, поставьте под свет в помещении и дождитесь награды. Этот вид нарушает однообразие прямостоячих растений и особенно привлекателен, придавая движение склону холма.
Закатный папоротник оправдывает свое название
Название: Dryopteris lepidopoda
Зоны: от 6 до 9
Размер: от 15 до 24 дюймов в высоту и 2 фута в ширину
Условия: светлый оттенок; богатая гумусом влажная, но не влажная почва
Общие названия различаются по полезности, но это прозвище – идеальный выбор для этого потрясающего и адаптируемого вида с его теплыми розовыми / лососевыми / красными / желтыми весенними порослями в форме вазы.В течение всего лета продолжают выращиваться новые красочные листья, которые непрерывно демонстрируют и сияют слегка затененным участкам. Закатный папоротник с его неизменно вечнозеленым лицом служит элегантным обрамлением, подчеркивающим лесную тропинку, или манящим центром, привлекающим людей в сад. Он напоминает разноцветный древесный папоротник Валлиха (Dryopteris wallichiana, зоны 5–7), но меньше ростом и немного более устойчив к холоду. Относительно новый в выращивании, его может быть трудно найти на рынке, но он стоит всех усилий, даже для энтузиаста, не связанного с папоротниками.
Перевернутый папоротник образует перистую вазу
Название: Arachniodes standishii
Зоны: от 5 до 9
Размер: от 2 до 3 футов в высоту и 2 фута в ширину
Условия: От частичной до полной тени; влажная рыхлая органическая почва
Перевернутый папоротник образует прочную вазу из кружевной листвы с тонким узором светлого оттенка. Видные прожилки на верхушке листьев дали этому папоротнику название. Есть два типа: Корейская версия несколько больше японской и является наиболее распространенной в культуре.Он составляет привлекательный контраст с более тяжелыми вечнозелеными деревьями.
Азиатский саблезубый папоротник противостоит снегу
Название: Polystichum neolobatum
Зоны: 5-8
Размер: от 18 до 30 дюймов в высоту и 2 фута в ширину
Условия: от светлого до глубокого оттенка; влажная почва
Азиатский саблезубый папоротник не только вечнозеленый, но и один из немногих папоротников, чьи листья сохраняют свой вертикальный вид даже после того, как их закопают в снег на фут или около того. Вайи узкие, блестящие, на ощупь заметно колючие.Новый весенний рост вызывает интерес благодаря скоплениям фиддлхэдов, густо покрытых декоративными красновато-коричневыми чешуйками. Он горит на солнце и не любит пересыхать, но в остальном этот папоротник можно расположить практически в любой глубине тени.
Они почти заботятся о себе
Хотя папоротники не требуют особого ухода, они не лишены нужды. Как выращивающему папоротник мне снова и снова задают одни и те же вопросы:
Когда сажать папоротники?
Папоротники можно сажать всякий раз, когда почва не промерзла, но лучше всего ранней осенью, чтобы они успели укорениться до зимнего стресса.Обязательным условием является поддержание влажности папоротников, пока они адаптируются к новому дому и приживаются их корни. Даже засухоустойчивые растения нуждаются в воде в течение первого лета.
Нужно ли их удобрять?
Папоротники следует высаживать в рыхлую смесь компоста с хорошим дренажем. При правильной настройке удобрения не нужны, хотя я позволяю опавшим листьям деревьев обогащать почву, оставаясь мульчей вокруг растений. И никогда не добавляйте лайм в папоротниковый сад.
Могу я их разделить?
Папоротники со стелющимися или множественными венчиками (фото вверху) являются кандидатами на деление. Идеальное время – осень, чтобы дать трансплантатам время прижиться без стресса от тепла и до стресса от холода. Острой лопатой аккуратно срежьте потомство. Пересаживайте членики в хорошую почву и полутень и держите их поливными.
Следует ли обрезать листья весной?
Листопадные папоротники ухаживают за собой и осенью откладывают листья.Тем не менее, листья вечнозеленого папоротника будут продолжать давать пищу всю зиму, поэтому лучше оставить их в покое до того момента, когда они начнут новый рост весной. Затем их можно срезать, но будьте осторожны, чтобы не срезать новые листья.
Есть ли у них вредители?
Папоротники, к счастью, не имеют проблем с вредителями. Слизняки и улитки сочтут уязвимые новообразования аппетитными, и их следует избегать с помощью раннего применения приманки для слизняков. Распределите приманку на расстоянии, чтобы не привлекать к своим лакомствам хищников.Тля редка, но, опять же, ее привлекают нежные новообразования. Их можно отразить легкой струей воды из шланга. Олени обычно не беспокоят папоротники; однако кролики считают папоротники и все остальное в саду своими личными салат-барами.
Чилийский твердый папоротник – архитектурное чудо
Название: Blechnum chilense
Зоны: с 8 по 11
Размер: от 3 до 5 футов в высоту, неограниченное распространение
Условия: От частичного до глубокого оттенка; влажная, хорошо дренированная почва
Чилийцы называют этот папоротник las costillas de la vaca, или коровьи ребра – подходящее название.Чилийский твердый папоротник обладает великолепной архитектурой. Его светящиеся красные новообразования (которые становятся более заметными, если папоротник не удобрять и не на богатых питательными веществами почвах) превращаются в идущие вверх статные стебли листьев. Он выдержит слегка сухой оттенок, но не будет таким ярким. А с его ползучими корневищами он действительно может быть крепким, когда счастлив (настолько, что невозможно определить ширину). Он будет распространяться с энтузиазмом, но не агрессивен или агрессивен.
Древесный папоротник Зибольда на самом деле папоротник
Название: Dryopteris sieboldii
Зоны: от 6 до 9
Размер: 30 дюймов в высоту и ширину
Условия: светлый оттенок; богатая гумусом влажная почва
Древесный папоротник Зибольда – это растение с выдающимся и уникальным очертанием, которое побуждает некоторых посетителей спрашивать: «Это действительно папоротник?» Это смелая и очень декоративная тема для разговора, которая особенно хороша, когда ее поощряет теплота.В более прохладных местах он разрастается довольно поздно в сезоне и несколько экономно производит эффектную новую листву. Вайи выглядят как длинные кинжалы и имеют пастельный сине-зеленый цвет – особенно привлекательны, если их посадить среди коряги или замшелых бревен, или когда они подкреплены светлой или очень темной лиственной фольгой.
Папоротник падуба Фортуны такой же крепкий, как и жирный
Название: Cyrtomium fortunei
Зоны: 5-10
Размер: 15 дюймов в высоту и 18 дюймов в ширину
Условия: От частичной до полной тени; плодородная, гумусовая, влажная, хорошо дренированная почва
Папоротник падуба Fortune – самый холодостойкий из всех папоротников падуба (Cyrtomium spp., Зоны 5–11). Намек на падуб относится к необычной форме ушных раковин папоротника, которые при некотором воображении напоминают листья падуба. Это придает роду внешний вид, сильно отличающийся от стереотипного папоротника с кружевными листьями, и делает этот вид отличным выбором для создания интересного контраста с изящным внешним видом более традиционных папоротников. Это отличный выбор для глубокого оттенка, где его бледно-зеленые вечнозеленые лиственные розетки привносят легкость в окружающую среду.Как и многие другие папоротники, он служит прекрасной пленкой для цветущих весной цветов. А если поместить его вдали от солнца, он также станет простым в уходе комнатным растением.
Гималайский папоротник девичьих волос образует оборчатый ковер
Название: Adiantum venustum
Зоны: 5 (с охраной) до 9
Размер: от 12 до 15 дюймов, ползать бесконечно
Условия: полутень; влажная, хорошо дренированная почва
В отличие от более знакомых нам местных папоротников девичьей шерсти, образующих комковицы (Adiantum aleuticum и A.pedatum, зоны 3–8), этот вид будет ползать медленно, но не агрессивно, образуя элегантный вечнозеленый ковер с мелкой текстурой. Несмотря на свой нежный вид, гималайский папоротник девичьей шерсти не требователен к почве и среде обитания, но, как и большинство других папоротников, он процветает в отфильтрованной тени и на компостированной почве. Весной новые листья сначала появляются в виде бледно-розовых усиков, которые медленно разворачиваются и обычно не страдают от поздних заморозков. Хотя это не является необходимым для здоровья растения, старые листья можно обрезать до весны, чтобы привести растение в порядок.Этот папоротник можно разделить осенью, но маленькие кусочки плохо восстанавливаются, поэтому делайте значительные части.
Источники
Следующие продавцы растений с доставкой по почте предлагают самый широкий выбор представленных папоротников:
Fancy Fronds Nursery, золотой слиток, штат Вашингтон .; 360-793-1472; fancyfrondsnursery.com
Foliage Gardens, Bellevue, Вашингтон .; 425-747-2998; foliagegardens.com
Питомник растительных наслаждений, Роли, Н.C .; 919-772-4794; plantdelights.com
Sebright Gardens, Салем, штат Орегон; 503-463-9615; sebrightgardens.com
Сью Олсен – владелица Лиственных садов в Белвью, Вашингтон, и автор Энциклопедии садовых папоротников.
Фотографии, если не указано иное: любезно предоставлены Сью Олсен; п. 44, millettephotomedia.com; п. 45, любезно предоставлено Ричи Штеффеном / Great Plant Picks; п. 46 (вверху), любезно предоставлено Casa Flora; п. 48 (вверху), любезно предоставлено Sebright Gardens; п. 49, Мишель Жерве
Растения папоротников на продажу | Купить большие папоротники онлайн
Папоротниковые растения
Благодаря своим листьям и кружевным листьям папоротники прекрасно впишутся в ландшафтный сад и станут комплиментом деревьям, цветущим кустарникам и другим уличным растениям.Около 12 000 видов папоротников произрастают по всему миру в различных местообитаниях. Многие подходят для сада. Бостонский папоротник, папоротник оленьего рога и другие могут жить даже в помещении. Существует множество разновидностей папоротника, которые сделают ваш ландшафт более выразительным и интересным. Папоротники выглядят потрясающе, посаженные за другими небольшими растениями, такими как Variegated Hosta. Если посадить достаточно полно и густо, они создадут самую красивую заднюю часть. Купите наши самые продаваемые папоротники: Lady Fern, Hay Scented Fern и Ostrich Fern.
Каждая разновидность папоротника растет в пятнистом свете, создаваемом ветвями деревьев, как они растут в лесу. Постарайтесь смоделировать эти условия, когда будете сажать их в своем ландшафте. Учтите, что чем больше прямых солнечных лучей попадает на папоротник, тем больше влаги ему требуется. Однако растения папоротника могут пережить короткие засушливые периоды, что делает их особенно простыми в уходе. Мульчируйте вокруг папоротника мелко измельченные листья или листовую плесень.Это будет имитировать состояние естественной лесной подстилки. Добавление слоя каждую весну поможет им удерживать влагу в летнюю жару.
Так как папоротник – это растение с мелкими корнями, тщательно пропалывайте его сорняками, наблюдая за прорастающими грызунами. Вам не нужно беспокоиться о дополнительных удобрениях для папоротника. Компост и мульча из листовой плесени обеспечивают все необходимые питательные вещества.