404 Cтраница не найдена
Размер:
A
A
A
Цвет: CCC
Изображения Вкл.Выкл.
Обычная версия сайта
RU EN
Башкирский государственный медицинский университет
- Университет
- Руководство
- Ректорат
- Обращение к ректору
- Ученый совет
- Университету 90 лет
- Телефонный справочник
- Документы
- Структура
- СМИ о вузе
- Символика БГМУ
- Электронный ящик доверия
- Комплексная программа развития БГМУ
- Антитеррор
- Сведения об образовательной организации
- Абитуриенту
- Обращение граждан
- Фотогалерея
- Карта сайта
- Видеогалерея
- Доступная среда
- Оплата банковской картой
- Реорганизация вуза
- Календарь мероприятий
- Образование
- Учебно-методическое управление
- Организация учебного процесса в условиях предупреждения Covid-19
- Учебно-организационный отдел
- Центр практических навыков
- Факультеты
- Кафедры
- Институт дополнительного профессионального образования
- Медицинский колледж
- Деканат по работе с иностранными обучающимися
- Управление международной деятельности
- Отдел ординатуры
- Расписание
- Менеджмент качества
- Федеральный аккредитационный центр
- Научно-образовательный медицинский кластер «Нижневолжский»
- Государственная итоговая аттестация
- Первичная аккредитация
- Первичная специализированная аккредитация
- Внутренняя оценка качества образования
- Информация для инвалидов и лиц с ограниченными возможностями здоровья
- Информация для студентов
- Я-профессионал
- Всероссийская студенческая олимпиада по хирургии с международным участием
- Медицинский инспектор
- Онлайн обучение
- Социальная работа в системе здравоохранения
- Новые образовательные программы
- Электронная учебная библиотека
- Независимая оценка качества образования
- Наука и инновации
- Наука и университеты
- Структура и документы
- Указ Президента Российской Федерации «О стратегии научно-технологического развития Российской Федерации»
- Стратегия развития медицинской науки до 2025 года
- Научно-исследовательские подразделения
- Клинические исследования и испытания, ЛЭК
- Диссертационные советы
- Докторантура
- Аспирантура
- Грантовая политика БГМУ
- Актуальные гранты, стипендии, конкурсы
- Конференции и форумы
- Гранты, премии, конкурсы, конференции для молодых ученых
- Полезные интернет-ссылки
- Научные издания
- Проблемные научные комиссии
- Патентная деятельность
- БГМУ в рейтингах университетов
- Публикационная активность
- НИИ кардиологии
- Биобанк
- Репозиторий БГМУ
- Евразийский НОЦ
- Лечебная работа
- Клиника БГМУ
- Всероссийский центр глазной и пластической хирургии
- Клиническая стоматологическая поликлиника
- Клинические базы
- Отчеты по лечебной работе
- Договорная работа с клиническими базами
- Отделения клиники БГМУ
- Лицензии
- Санаторий-профилакторий БГМУ
- Жизнь БГМУ
- Воспитательная и социальная работа
- Отдел по культурно-массовой работе
- Отдел по связям с общественностью
- Общественные объединения и органы самоуправления
- Отдел по воспитательной и социальной работе
- Творческая жизнь
- Спортивная жизнь
- Профсоюз обучающихся БГМУ
- Совет кураторов
- Ассоциация выпускников
- Работа музеев на кафедрах
- Выпускники БГМУ – ветераны ВОВ
- Золотой фонд БГМУ
- Медиа центр
- БГМУ – ВУЗ здорового образа жизни
- Юбиляры
- Жизнь иностранных студентов БГМУ
- Университету 90 лет
- Университету 85 лет
- Празднование 75-летия Победы в Великой Отечественной войне
- Научная библиотека
- Приоритет 2030
- О программе
- Проектный офис
- Стратегические проекты
- Миссия и стратегия
- Цифровая кафедра
- Конкурсы для студентов
- Отчетность
- Публикации в СМИ
- Программа развития
- Научные семинары для студентов и ученых БГМУ
- Главная
Vzaimodejstvie Genov : Основы генетики : Все про гены!
Взаимодействие аллельных генов
В состав генотипа входит большое количество генов, функционирующих и взаимодействуют как целостная система. Г. Мендель в своих опытах обнаружил только одну форму взаимодействия между аллельными генами – полное доминирование одной аллели и полную рецесивнисть другой. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Г. Мендель в своих опытах обнаружил только одну форму взаимодействия между аллельными генами – полное доминирование одной аллели и полную рецесивнисть другой. Генотип организма нельзя рассматривать как простую сумму независимых генов, каждый из которых функционирует вне связи с другими. Фенотипное проявления того или иного признака являются результатом взаимодействия многих генов.
Различают две основных группы взаимодействия генов: взаимодействие между аллельными генами и взаимодействие между неаллельнимы генами. Однако следует понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме происходит взаимодействие между белками – ферментами, синтез которых опрелятся генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1) для образования определенного признака необходимо взаимодействие двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2) фермент, что был синтезирован с участием одного гена, полностью подавляет или инактивирует действие фермента, что был образован другим неаллельным геном;
3) два ферменты, образование которых контролируется двумя неаллельми генами, влияющими на один признак или на один процесс так, что их совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов. Гены, которые занимают идентичные (гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия – полное доминирование, которое впервые описано Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление одной из аллелей доминирует над проявлением другой. При полном доминировании расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу – 3:1. В медицинской практике с двух тысяч моногенных наследственных болезней почти в половины имеет место доминированое проявления патологических генов над нормальными. В гетерозигот патологический аллель проявляется в большинстве случаев признаками заболевания (доминантный фенотип).
Неполное доминирование – форма взаимодействия, при которой у гетерозиготного организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а), вследствие чего проявляется промежуточный между родительскими признак. Здесь расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов вызывает формирование зависимого от него продукта, то есть оказываются продукты обеих аллелей. Классическим примером такого проявления является система групп крови, в частности система АBО, когда эритроциты человека несут на поверхности антигены, контролируемые обеими аллелями. Такая форма проявления носит название кодоминированием.
Сверхдоминирование – когда доминантный ген в гетерозиготном состоянии проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе АА-нормальная продолжительность жизни; Аа – удлиненная триватисть жизни; аа – летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в природе количество аллелей может быть более двух, если какой то локус может находится в разных состояниях. В таких случаях говорят о множественные аллели или множественный аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами, например: А, А1, А3 … Аллельные гена локализуются в одинаковых участках гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с различием гомологичних хромосом) попадает только по одному из них. Для множественных аллелей характерное влияние всех аллелей на один и тот же признак. Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивный относительно первого, однако доминирует над следующими и т.д. Одним из примеров проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение, поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими продукты. При этом имеет место отклонение от менделивских закономерностей расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность, эпистаз, полимерию и модифицирующее действие (плейотропия).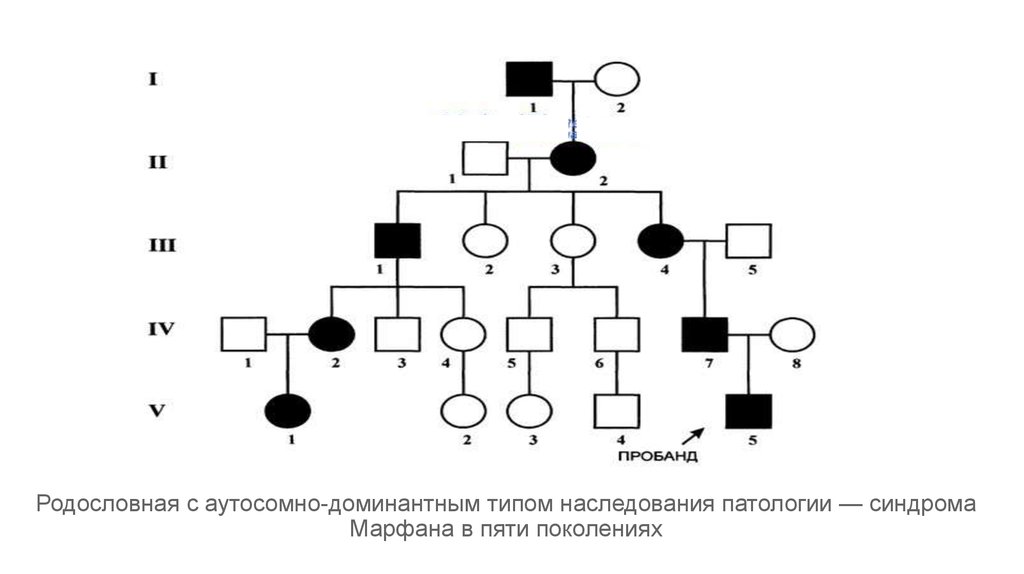
Комплементарность это такой тип взаимодействия неаллельних генов, когда один доминантный ген дополняет действие другого неаллельного доминантного гена, и они вместе определяют новый признак, который отсутствует у родителей. Причем соответственный признак развивается только в присутствии обоих неаллельних генов. Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа) – альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез защитного белка – интерферона. Его образование в организме связано с комплементарным взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз -это такое взаимодействие неаллельных генов, при котором один ген подавляет действие другого неаллельного гена. Угнетение могут вызывать как доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого розличают эпистаз доминантный и рецессивный. Подавляющий ген получил название ингибитора или супрессора. Гены-ингибиторы в основном не детерминируют развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3; 12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом взаимодействия.
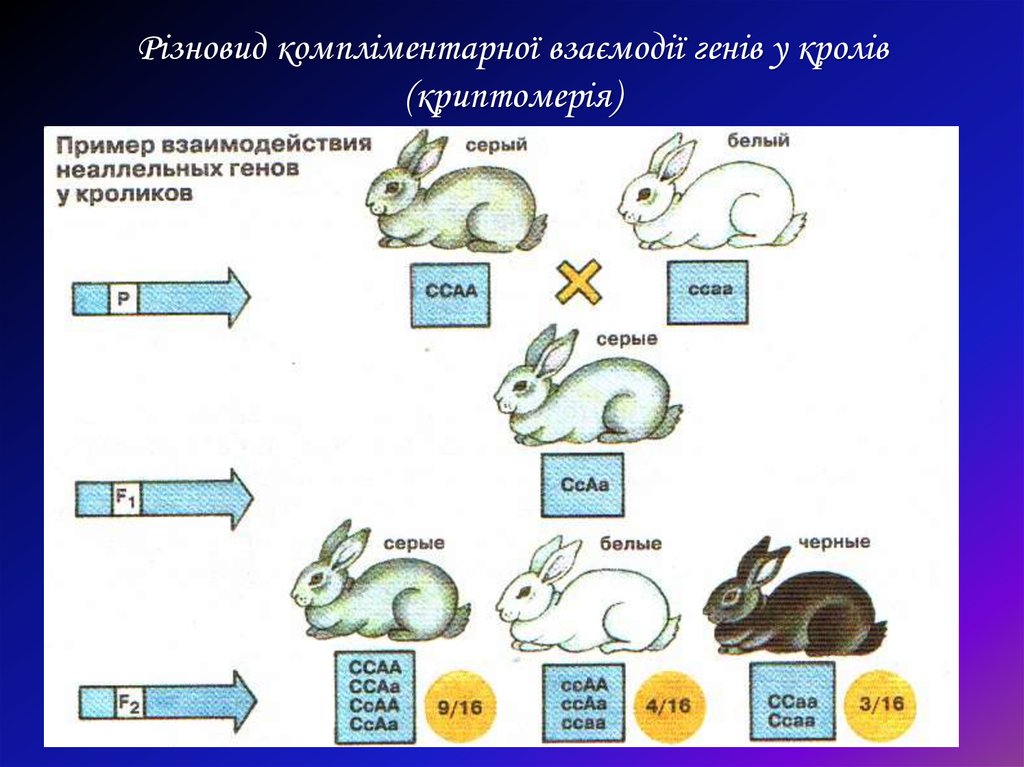
Если ген-супрессор рецессивный, то возникает криптомерия (греч. хриштад – тайный, скрытый). У человека таким примером может быть “Бомбейский феномен”. В этом случае редкий рецессивный аллель “х” в гомозиготном состоянии (мм) подавляет активность гена jB (определяющий В (III) группу крови системы АВО).![]() Поэтому женщина с генотипом jв_хх, фенотипно имеет I группу крови – 0 (I).
Поэтому женщина с генотипом jв_хх, фенотипно имеет I группу крови – 0 (I).
Полигенное наследования количественных признаков
– плейотропия
– экспрессивность и пенетрантность генов
Большинство количественных признаков организмов определяется несколькими неаллельнимы генами (полигенами). Взаимодействие таких генов в процессе формирования признака называется полимерным. В этом случае две или более доминантных аллели в равной степени влияют на развитие одной и того же признаки. Поэтому полимерные гены принято обозначать одной буквой латинского алфавита с цифровым индексом, например: А1А1 и а1а1. Впервые однозначные факторы были выявлены шведским генетиком Нильсон-Эле (1908 г.) при изучении наследования цвета в пшеницы. Было установлено, что этот признак зависит от двух полимерных генов, поэтому при скрещивании доминантних и рецессивных дигомозигот – окрашенной (А1А1, А2 А2) с бесцветной (а1а1, а2а2) – в F, все растения дают окрашенные семена, хотя они светлее, чем родительские экземпляры, которые имеют красное семя. В F, при скрещивании особей первого поколения проявляется расщепление по фенотипу в соотношении 15:1, потому бесцветным является лишь рецессивные дигомозиготы (а1а1 а2а2). В пигментированных экземплярах интенсивность цвета очень отличается в зависимости от количества полученных ими доминантних аллелей: максимальная в доминантных дигомозигот (А1А1, А2 А2) и минимальная у носителей одного из доминантных аллелей.
В F, при скрещивании особей первого поколения проявляется расщепление по фенотипу в соотношении 15:1, потому бесцветным является лишь рецессивные дигомозиготы (а1а1 а2а2). В пигментированных экземплярах интенсивность цвета очень отличается в зависимости от количества полученных ими доминантних аллелей: максимальная в доминантных дигомозигот (А1А1, А2 А2) и минимальная у носителей одного из доминантных аллелей.
Важная особенность полимерии – суммация действия неаллельних генов на развитие количественных признаков. Если при моногенном наследовании признака возможны три варианта “доз” гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до четырех и более. Суммация “доз” полимерных генов обесчивает существования непрерывных рядов количественных изменений.
Биологическое значение полимерии заключается еще и в том, что признаки, кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном. Организм без полимерных генов был бы очень неустойчивым: любая мутация или рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет неблагоприятный характер.
У животных и растений есть много полигенных признаков, среди них и ценные для хозяйства: интенсивность роста, скороспелость, яйценоскость, количество молока, содержание сахаристых веществ и витаминов и т.п.
Пигментация кожи у человека определяется пятью или шестью полимерными генами. В коренных жителей Африки (негроидной расы) преобладают доминантные аллели, у представителей европеоидной расы – рецессивные. Поэтому мулаты имеют промежуточную пигментацию, но при браках мулатов у них возможно появление как более, так и менее интенсивно пигментированных детей.
Многие морфологические, физиологические и патологические особенности человека определяются полимерными генами: рост, масса тела, величина артериального давления и др. Развитие таких признаков у человека подчиняется общим законам полигенного наследования и зависит от условий среды. В этих в случаях наблюдается, например, склонность к гипертонической болезни, ожирению и др. Данные признаки при благоприятных условиях среды могут не проявиться или проявиться незначительно. Эти полигенные признаки отличаются от моногенных. Изменяя условия среды можно обеспечить профилактику ряда полигенных заболеваний.
Эти полигенные признаки отличаются от моногенных. Изменяя условия среды можно обеспечить профилактику ряда полигенных заболеваний.
Плейотропия
Плейотропное действие генов – это зависимость нескольких признаков от одного гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает плодовитость, уменьшает продолжительность жизни. У человека известна наследственная болезнь – арахнодактилия (“паучьи пальцы”-очень тонкие и длинные пальцы), или болезнь Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной ткани и одновременно влияет на развитие нескольких признаков: нарушение строения хрусталика глаза, аномалии в сердечно-сосудистой системе.
Плейотропное действие гена может быть первичным и вторичным. При первичной плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
При этом поражаются одновременно мембраны эпителиальных клеток кишечника и почечных канальцев с расстройствами пищеварительной и выделительной систем.
При вторичной плейотропии есть один первичный фенотипний проявление гена, вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к множественным эффектам. Так, при серповидно клеточной анемии у гомозигот наблюдается несколько патологических признаков: анемия, увеличенная селезенка, поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные проявления гена составляют иерархию вторичных проявлений. Первопричиной, непосредственным фенотипним проявлением дефектного гена является аномальный гемоглобин и эритроциты серповидной формы. Вследствие этого происходят последовательно другие патологические процессы: слипание и разрушение эритроцитов, анемия, дефекты в почках, сердце, мозге – эти патологические признаки вторичны.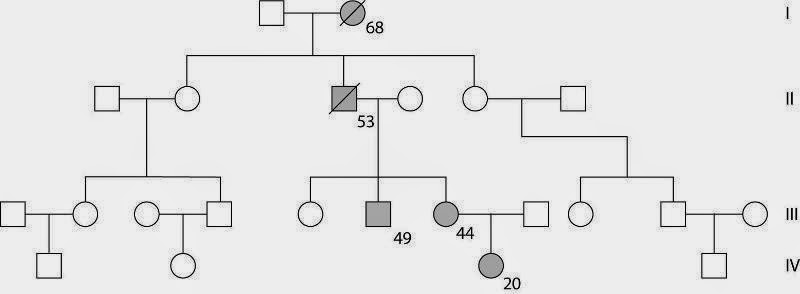
При плейотропии, ген, воздействуя на какой то один основнй признак, может также менять, модифицировать проявление других генов, в связи с чем введено понятие о генах-модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых “основным” геном.
Показателями зависимости функционирования наследственных задатков от характеристик генотипа является пенетрантность и экспрессивность.
Рассматривая действие генов, их аллелей необходимо учитывать и модифицирующее влияние среды, в которой розвивается организм. Если растения примулы скрещивать при температуре 15-20 ° С, то в F1 согласно менделивской схеме, все поколения будут иметь розовые цветы. Но когда такое скрещивание проводить при температуре 35 °С, то все гибриды будут иметь цветы белого цвета. Если же осуществлять скрещивания при температуре около 30 ° С, то возникает разное соотношение (от 3:1 до 100%) растений с белыми цветами.
Такое колебание классов при расщеплении в зависимости от условий среды получило название пенетрантность – сила фенотипного проявления. Итак, пенетрантность – это частота проявления гена, явление появления или отсутствия признака у организмов, одинаковых по генотипу.
Итак, пенетрантность – это частота проявления гена, явление появления или отсутствия признака у организмов, одинаковых по генотипу.
Пенетрантность значительно колеблется как среди доминантных, так и среди рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании определенных условий и достаточно редких внешних условий (высокая пенетрантность), у человека есть гены, фенотипное проявление которых происходит при любых соединениях внешних условий (низкая пенетрантность). Пенетрантностью измеряется процентом организмов с фенотипным признаком от общего количества обследованных носителей соответствующих аллелей.
Если ген полностью, независимо от окружающей среды, определяет фенотипное проявление, то он имеет пенетрантность 100 процентов. Однако некоторые доминантные гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное наследования, но бывают пропуски поколений. Доминантная аномалия – преждевременное половое созревание – присуще только мужчинам, однако иногда может передаться заболевания от человека, который не страдал этой патологией. Пенетрантностью указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды.
Пенетрантностью указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак, пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не константное свойство гена, а функция генов в конкретных условиях среды.
Экспрессивность (лат. ехргеssio – выражение) – это изменение количественного проявления признака в разных особей-носителей соответствующего аллелей.
При доминантных наследственных заболеваниях экспрессивность может колебаться. В одной и той же рсемье могут проявляться наследственные болезни от легких, едва заметных до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д. Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и имеют незначительные колебанийния экспрессивности.
Генетическая дифференциация и эволюционная адаптация Cryptomeria japonica
. 2014 14 октября; 4 (12): 2389-402.
дои: 10. 1534/g3.114.013896.
1534/g3.114.013896.
Ёсихико Цумура 1 , Кентаро Утияма 2 , Ёсинари Моригути 3 , Мегуми К Кимура 2 , Санеоши Уэно 2 , Токуко Уджино-Ихара 2
Принадлежности
- 1 Департамент генетики леса, Научно-исследовательский институт лесного хозяйства и лесных товаров, Цукуба Ибараки 305-8687, Япония [email protected].
- 2 Департамент генетики леса, Научно-исследовательский институт лесного хозяйства и лесных товаров, Цукуба Ибараки 305-8687, Япония.
- 3 Высшая школа науки и технологий, Университет Ниигата, 8050, Игараши 2-Ночо, Ниси-ку Ниигата 950-2181, Япония.

- PMID: 25320072
- PMCID: PMC4267934
- DOI: 10.1534/г3.114.013896
Бесплатная статья ЧВК
Ёсихико Цумура и др. G3 (Бетесда). .
Бесплатная статья ЧВК
. 2014 14 октября; 4 (12): 2389-402.
дои: 10.1534/g3.114.013896.
Авторы
Ёсихико Цумура 1 , Кентаро Утияма 2 , Ёсинари Моригути 3 , Мегуми К Кимура 2 , Санеоши Уэно 2 , Токуко Уджино-Ихара 2
Принадлежности
- 1 Департамент генетики леса, Научно-исследовательский институт лесного хозяйства и лесных товаров, Цукуба Ибараки 305-8687, Япония ytsumu@ffpri. affrc.go.jp.
- 2 Департамент генетики леса, Научно-исследовательский институт лесного хозяйства и лесных товаров, Цукуба Ибараки 305-8687, Япония.
- 3 Высшая школа науки и технологий, Университет Ниигата, 8050, Игараши 2-Ночо, Ниси-ку Ниигата 950-2181, Япония.
 affrc.go.jp.
affrc.go.jp.- PMID: 25320072
- PMCID: PMC4267934
- DOI: 10.1534/г3.114.013896
Абстрактный
Локальная адаптация видов растений является центральной проблемой для выживания во время глобального изменения климата, особенно для долгоживущих лесных деревьев с их длительным временем регенерации и пространственно ограниченным потоком генов. Идентификация локусов и/или геномных регионов, связанных с местной адаптацией, необходима для получения знаний как об эволюции, так и о молекулярном размножении в связи с изменением климата. Cryptomeria japonica – важный вид для лесного хозяйства Японии; он имеет широкое естественное распространение и может выжить в различных средах. Генетическая структура 14 природных популяций этого вида исследована с использованием 3930 маркеров SNP. Население со стороны Тихого океана в Японии явно отличается от населения со стороны Японского моря, как обсуждалось в предыдущих исследованиях. Структурный анализ и деревья сети популяций показывают, что периферийные популяции, в том числе самые северные и южные, обладают уникальными особенностями. Мы обнаружили, что коэффициент генетической дифференциации низок, FST = 0,05, хотя он должен учитывать наличие важных генов, связанных с адаптацией к специфическим условиям окружающей среды. Всего было обнаружено 208 локусов-выбросов, из которых 43 были связаны с переменными среды.
Идентификация локусов и/или геномных регионов, связанных с местной адаптацией, необходима для получения знаний как об эволюции, так и о молекулярном размножении в связи с изменением климата. Cryptomeria japonica – важный вид для лесного хозяйства Японии; он имеет широкое естественное распространение и может выжить в различных средах. Генетическая структура 14 природных популяций этого вида исследована с использованием 3930 маркеров SNP. Население со стороны Тихого океана в Японии явно отличается от населения со стороны Японского моря, как обсуждалось в предыдущих исследованиях. Структурный анализ и деревья сети популяций показывают, что периферийные популяции, в том числе самые северные и южные, обладают уникальными особенностями. Мы обнаружили, что коэффициент генетической дифференциации низок, FST = 0,05, хотя он должен учитывать наличие важных генов, связанных с адаптацией к специфическим условиям окружающей среды. Всего было обнаружено 208 локусов-выбросов, из которых 43 были связаны с переменными среды. С помощью анализа сцепления в геноме были обнаружены четыре сгруппированные области локусов-выбросов. Неравновесие по сцеплению (LD) было довольно высоким в этих скоплениях выделенных локусов, которые были обнаружены в группах сцепления (LGs) 2, 7, 10 и 11, особенно между популяциями двух разновидностей, а также при обнаружении межхромосомного LD. Регион LG7 характерен для популяции Якусима, которая представляет собой большую изолированную периферийную популяцию, занимающую специфическую среду в результате изоляции в сочетании с вулканической активностью в регионе. Обнаруженная LD может служить убедительным доказательством отбора между разновидностями.
С помощью анализа сцепления в геноме были обнаружены четыре сгруппированные области локусов-выбросов. Неравновесие по сцеплению (LD) было довольно высоким в этих скоплениях выделенных локусов, которые были обнаружены в группах сцепления (LGs) 2, 7, 10 и 11, особенно между популяциями двух разновидностей, а также при обнаружении межхромосомного LD. Регион LG7 характерен для популяции Якусима, которая представляет собой большую изолированную периферийную популяцию, занимающую специфическую среду в результате изоляции в сочетании с вулканической активностью в регионе. Обнаруженная LD может служить убедительным доказательством отбора между разновидностями.
Ключевые слова: ЛД; хвойное дерево; местная адаптация; место выброса; выбор.
Авторское право © 2014 Tsumura et al.
Цифры
Рисунок 1
Естественное распространение Cryptomeria japonica…
Рисунок 1
Естественное распространение Cryptomeria japonica в Японии (заштрихованные области) (Hayashi 1951) и…
Рисунок 1 Естественное распространение Cryptomeria japonica в Японии (заштрихованные области) (Hayashi 1951) и местонахождение 14 природных популяций, обследованных в ходе этого исследования. Пунктирная линия указывает на береговую линию примерно 18 000 лет назад. Области, заштрихованные жирным шрифтом или обведенные тонкими диагональными линиями, указывают на установленные рефугиумы (полуостров Идзу, залив Вакаса, остров Оки и остров Якусима) и вероятные рефугиумы, соответственно, в то время (Цукада 19).86).
Пунктирная линия указывает на береговую линию примерно 18 000 лет назад. Области, заштрихованные жирным шрифтом или обведенные тонкими диагональными линиями, указывают на установленные рефугиумы (полуостров Идзу, залив Вакаса, остров Оки и остров Якусима) и вероятные рефугиумы, соответственно, в то время (Цукада 19).86).
Рисунок 2
Генетические связи между 14…
Рисунок 2
Генетические отношения среди 14 популяций, обследованных с помощью STRUCTURE (Pritchard et al. 2000)…
фигура 2 Генетические отношения среди 14 популяций, обследованных с помощью STRUCTURE (Pritchard и др. 2000) на основе 3930 SNP. Модели с K = 2 и K = 4 были оптимальными на основе значения дельта K и наивысшего значения логарифмического правдоподобия соответственно.
Рисунок 3
Сеть населения на основе…
Рисунок 3
Сеть населения, основанная на попарном анализе F ST значения между совокупностями, построенными…
Рисунок 3Сеть населения, основанная на парных F ST значениях между популяциями, построенными Neighbor-net.
Рисунок 4
График значений F ST …
Рисунок 4
График значений F ST и коэффициентов Байеса (log 10 ) для 3930…
Рисунок 4 График значений F ST и коэффициентов Байеса (log 10 ) для 3930 локусов, идентифицированных с использованием теста на выбросы BayeScan. Пунктирные линии показывают log 10 порога байесовского фактора, который дает «решающее» свидетельство выбора, соответствующее апостериорной вероятности 0,9.9.
Пунктирные линии показывают log 10 порога байесовского фактора, который дает «решающее» свидетельство выбора, соответствующее апостериорной вероятности 0,9.9.
Рисунок 5
Распределение выявленных маркеров…
Рисунок 5
Распределение маркеров, идентифицированных как потенциально вовлеченные в местную адаптацию, по…
Рисунок 5 Распределение маркеров, идентифицированных как потенциально вовлеченные в локальную адаптацию, в геноме Криптомерия японская . На рисунке показаны графики положения маркера (на основе 1255 картированных SNP или других маркеров ДНК; Moriguchi et al. 2012; Y. Moriguchi, T. Ujino-Ihara, K. Uchiyama, N. Futamura, S. Ueno, A. (неопубликованные данные Matsumoto и Y. Tsumura) в кумулятивных сантиморганах (сМ) по сравнению с (A) коэффициент Байеса (log 10 ), определенный с помощью анализа BayeScan, и (B) F – статистическое значение, Ф СТ . Вертикальные пунктирные линии разграничивают 11 групп сцепления. Четыре серые зоны обозначают области-кандидаты, связанные с местной адаптацией.
Tsumura) в кумулятивных сантиморганах (сМ) по сравнению с (A) коэффициент Байеса (log 10 ), определенный с помощью анализа BayeScan, и (B) F – статистическое значение, Ф СТ . Вертикальные пунктирные линии разграничивают 11 групп сцепления. Четыре серые зоны обозначают области-кандидаты, связанные с местной адаптацией.
Рисунок 6
Тепловые карты неравновесия сцепления…
Рисунок 6
Тепловые карты значений неравновесного сцепления (LD) ( r 2 )…
Рисунок 6Тепловые карты значений неравновесия по сцеплению (LD) ( r 2 ) четырех областей, LG2, LG7, LG10 и LG11, и значения p .
См. это изображение и информацию об авторских правах в PMC
это изображение и информацию об авторских правах в PMC
Похожие статьи
Сканирование генома для выявления генетической структуры и адаптивных генов природных популяций Cryptomeria japonica.
Цумура Ю., Кадо Т., Такахаши Т., Тани Н., Уджино-Ихара Т., Ивата Х. Цумура Ю. и др. Генетика. 2007 август; 176 (4): 2393-403. doi: 10.1534/genetics.107.072652. Epub 2007 11 июня. Генетика. 2007. PMID: 17565947 Бесплатная статья ЧВК.
Сканирование генома для обнаружения адаптивных генов вдоль градиентов окружающей среды у японского хвойного дерева Cryptomeria japonica.
Цумура Ю., Учияма К., Моригучи Ю., Уэно С., Ихара-Уджино Т. Цумура Ю. и др. Наследственность (Эдинб).
2012 декабрь; 109 (6): 349-60. doi: 10.1038/hdy.2012.50. Epub 2012 29 августа.
Наследственность (Эдинб). 2012.
PMID: 22929151
Бесплатная статья ЧВК.Вывод демографической истории японского кедра Cryptomeria japonica с использованием секвенирования ампликонов.
Моригучи Н., Утияма К., Мияги Р., Морицука Э., Такахаши А., Тамура К., Цумура Ю., Тешима К.М., Тачида Х., Кусуми Дж. Моригути Н. и соавт. Наследственность (Эдинб). 2019 сен; 123 (3): 371-383. дои: 10.1038/s41437-019-0198-й. Epub 2019 26 февраля. Наследственность (Эдинб). 2019. PMID: 30809077 Бесплатная статья ЧВК.
Точное картирование генов мужской стерильности (MS1, MS2, MS3 и MS4) и разработка SNP-маркеров для селекции с помощью маркеров у кедра японского (Cryptomeria japonica D.
Don).Хасэгава Ю., Уэно С., Мацумото А., Уджино-Ихара Т., Учияма К., Тоцука С., Иваи Дж., Хакамата Т., Моригути Ю. Хасэгава Ю. и др. ПЛОС Один. 2018 15 ноября; 13 (11): e0206695. doi: 10.1371/journal.pone.0206695. Электронная коллекция 2018. ПЛОС Один. 2018. PMID: 30439978 Бесплатная статья ЧВК.
Поиск локусов под отбор: тренды, предубеждения и прогресс.
Аренс CW, Rymer PD, Stow A, Bragg J, Dillon S, Umbers KDL, Dudanec RY. Ahrens CW и соавт. Мол Экол. 2018 март; 27 (6): 1342-1356. doi: 10.1111/mec.14549. Epub 2018 30 марта. Мол Экол. 2018. PMID: 29524276 Обзор.
 2012 декабрь; 109 (6): 349-60. doi: 10.1038/hdy.2012.50. Epub 2012 29 августа.
Наследственность (Эдинб). 2012.
PMID: 22929151
Бесплатная статья ЧВК.
2012 декабрь; 109 (6): 349-60. doi: 10.1038/hdy.2012.50. Epub 2012 29 августа.
Наследственность (Эдинб). 2012.
PMID: 22929151
Бесплатная статья ЧВК.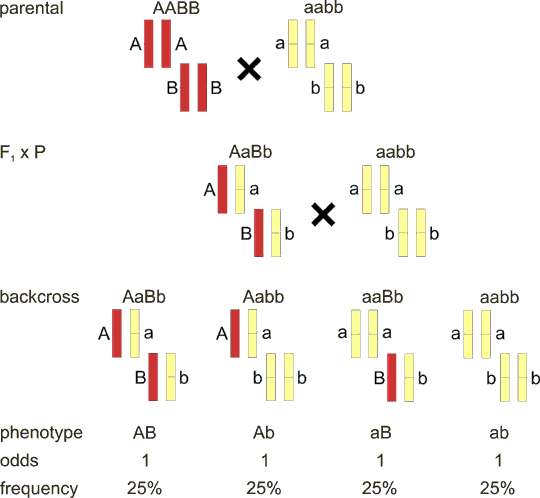 Don).
Don).Посмотреть все похожие статьи
Цитируется
Изменения годовой транскриптомной динамики клона кедра японского (Cryptomeria japonica D.
Don), высаженного в различных климатических условиях.Нос М., Ханаока С., Фукацу Э., Курита М., Миура М., Хираока Ю., Ики Т., Чигира О., Мисима К., Такахаши М., Ватанабэ А. Нос М. и др. ПЛОС Один. 2023 16 февраля; 18 (2): e0277797. doi: 10.1371/journal.pone.0277797. Электронная коллекция 2023. ПЛОС Один. 2023. PMID: 36795783 Бесплатная статья ЧВК.
Генетическое разнообразие и структура 4 -й цикла размножающейся популяции пихты китайской ( Cunninghamia lanceolata (lamb.) крючок).
Jing Y, Bian L, Zhang X, Zhao B, Zheng R, Su S, Ye D, Zheng X, El-Kassaby YA, Shi J. Цзин Ю и др. Фронт завод науч. 2023 27 января; 14:1106615. дои: 10.3389/fpls.2023.1106615. Электронная коллекция 2023. Фронт завод науч. 2023. PMID: 36778690 Бесплатная статья ЧВК.
Экологическая адаптация кутикулярных восков листьев наследуется для Medicago ruthenica .
Го И, Чжао С, Ли И, Ли З, Сяо Ц, Ван И, Чжан С, Ни Ю. Гуо Ю и др. Фронт завод науч. 2021 17 мая; 12:620245. doi: 10.3389/fpls.2021.620245. Электронная коллекция 2021. Фронт завод науч. 2021. PMID: 34079563 Бесплатная статья ЧВК.
Диверсификация выбросов терпеноидов предполагает географическую структуру, основанную на климате и составе патогенов японского кедра.
Хиура Т., Йошиока Х., Мацунага С.Н., Сайто Т., Кохяма Т.И., Кусумото Н., Утияма К., Суяма Ю., Цумура Ю. Хиура Т. и др. Научный представитель 2021 г. 15 апреля; 11 (1): 8307. doi: 10.1038/s41598-021-87810-x.
Научный представитель 2021.
PMID: 33859305
Бесплатная статья ЧВК.Идентификация и анализ генетического разнообразия гена мужской стерильности (MS1) у японского кедра (Cryptomeria japonica D. Don).
Хасэгава Ю., Уэно С., Вэй Ф.Дж., Мацумото А., Учияма К., Удзино-Ихара Т., Хакамата Т., Фуджино Т., Касахара М., Бино Т., Ямагути К., Сигенобу С., Цумура Ю., Моригути Ю. Хасэгава Ю. и др. Научный представитель 2021 15 января; 11 (1): 1496. doi: 10.1038/s41598-020-80688-1. Научный представитель 2021. PMID: 33452328 Бесплатная статья ЧВК.
 Don), высаженного в различных климатических условиях.
Don), высаженного в различных климатических условиях.
 Научный представитель 2021.
PMID: 33859305
Бесплатная статья ЧВК.
Научный представитель 2021.
PMID: 33859305
Бесплатная статья ЧВК.Просмотреть все статьи “Цитируется по”
Рекомендации
- Antao T., Lopes A., Lopes R., Beja-Pereira A., Luikart G., 2008. LOSITAN: инструмент для обнаружения молекулярной адаптации на основе метода Fst-выбросов. Биоинформатика BMC 9: 323.
–
ЧВК
–
пабмед
- Antao T., Lopes A., Lopes R., Beja-Pereira A., Luikart G., 2008. LOSITAN: инструмент для обнаружения молекулярной адаптации на основе метода Fst-выбросов.
- Аустерлиц Ф., Мариэтт С., Махон Н., Гуйон П. Х., Годель Б., 2000. Влияние процессов колонизации на генетическое разнообразие: различия между однолетними растениями и видами деревьев. Генетика 154: 1309–1321. – ЧВК – пабмед
- Бомонт М. А., Николс Р.А., 1996. Оценка локусов для использования в генетическом анализе структуры популяции. проц. биол. науч. 263: 1619–1626.
- Бомонт М.
- Бомонт М.А., Болдинг Д.Дж., 2004. Выявление адаптивной генетической дивергенции среди популяций на основе сканирования генома. Мол. Экол. 13: 969–980. – пабмед
- Богс Дж., Джаффе Ф.В., Такос А.М., Уокер А.Р., Робинсон С.П., 2007. Фактор транскрипции виноградной лозы VvMYBPA1 регулирует синтез проантоцианидина во время развития плодов. Завод Физиол. 143: 1347–1361. – ЧВК – пабмед
 Биоинформатика BMC 9: 323.
–
ЧВК
–
пабмед
Биоинформатика BMC 9: 323.
–
ЧВК
–
пабмед А., Николс Р.А., 1996. Оценка локусов для использования в генетическом анализе структуры популяции. проц. биол. науч. 263: 1619–1626.
А., Николс Р.А., 1996. Оценка локусов для использования в генетическом анализе структуры популяции. проц. биол. науч. 263: 1619–1626.Типы публикаций
термины MeSH
Организм | TreeGenes
Главная Каталог видов cryptomeria japonica
TreeGenes собирает геномную и феномную информацию для 2313 видов, представляющих 307 рода и 16 отрядов.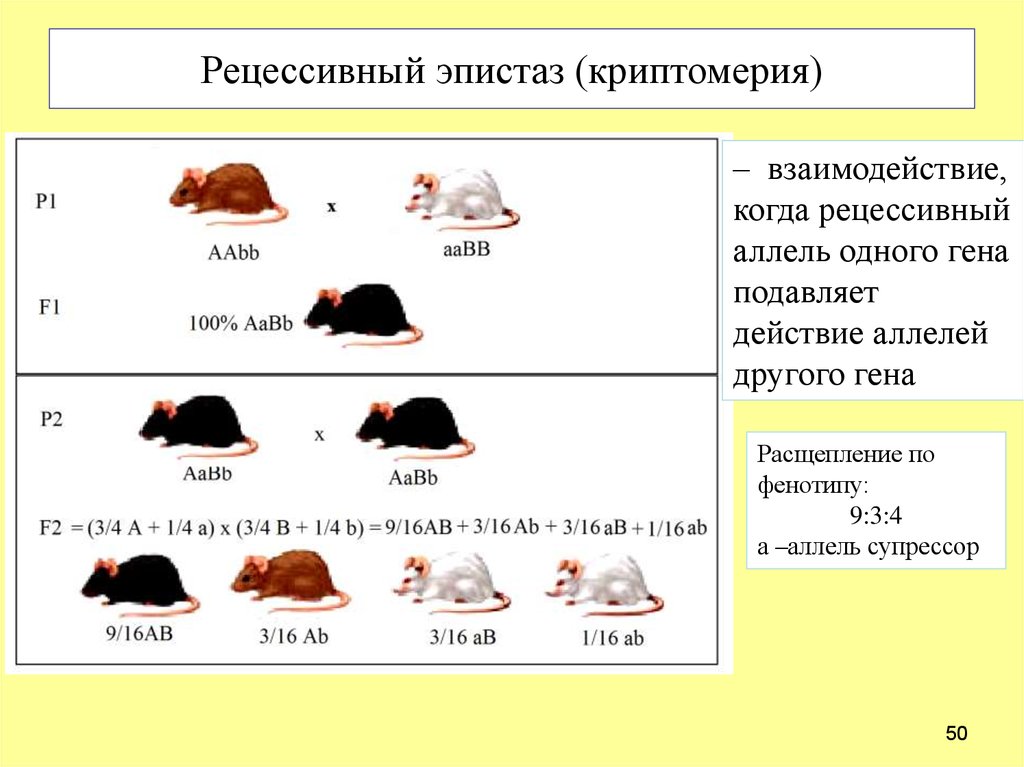 В этом разделе вы можете искать интересующие вас виды или просматривать ниже. Генерируются данные, связанные с каждым видом
из литературы, NCBI Genbank, PLAZA, а также из непосредственных материалов пользователей. В общем, виды включены, потому что мы имеем в
по крайней мере одна часть геномной или феномной информации, связанная с ними. Если вы хотите запросить добавление нового вида,
отправьте запрос
В этом разделе вы можете искать интересующие вас виды или просматривать ниже. Генерируются данные, связанные с каждым видом
из литературы, NCBI Genbank, PLAZA, а также из непосредственных материалов пользователей. В общем, виды включены, потому что мы имеем в
по крайней мере одна часть геномной или феномной информации, связанная с ними. Если вы хотите запросить добавление нового вида,
отправьте запрос
.
- Summary
- Transcriptome
- Literature
- PopGen
Species
cryptomeria japonica
Common Name
Japanese cedar
TreeGenes Code
Crja
Lineages
Viridiplantae; стрептофиты; стрептофитина; Эмбриофиты; трахеофиты; Эуфиллофита; Семенные растения; акрогимноспермы; пиниды; Кипарисовые; кипарисовые; Калоцедр
HAPLOID -хромосома номер
11
GBIF
, пытаясь загрузить информацию, это может занять несколько секунд
Potwo
, чтобы загрузить информацию, это может занять несколько секунд
.
32
NCBI EST
17917 EST | Скачать EST FASTA
кДНК NCBI
Скачать кДНК FASTA
NCBI TSA
9966 TSAs | Скачать ТСА ФАСТА 9
Последние публикации, связанные с японской криптомерией Saito, Maki, Sato, Masaya, Tsumura, Yoshihiko
Карта сцепления высокой плотности с 2560 маркерами и ее применение для локализации генов мужской стерильности ms3 и ms4 в cryptomeria japonica d. дон 2016
Определение стадий развития мужских стробилов с помощью цитологического анализа и анализа экспрессии генов у японского кедра (cryptomeria japonica) 2016; 36(5) 653-666
Инженерное микробное производство 2-пирона-4,6 -дикарбоновая кислота из остатков лигнина для использования в качестве химиката промышленной платформы 2016; 11(3) 6097-6109
 , Хаттори, Цутому, Миюки, Юрика, Эндо, Рёта
, Хаттори, Цутому, Миюки, Юрика, Эндо, Рёта Первое сообщение о гнили ствола, вызванной фомитипорией торрея, в префектуре Киото на сортах японского кедра, не связанных с ‘ санбу-суги 2016; 21(2) 105-109
Создание основной коллекции и оценка генетических ресурсов Cryptomeria japonica (кедр японский) 2015; 20(1) 186-196
Оценки наследуемости жесткости древесины и связанных с ней спектральных полос ближнего инфракрасного диапазона у клонов sugi (cryptomeria japonica) 2015; 20(1) 206-212
Карта сцепления высокой плотности с 2703 маркерами и ее применение для локализации генов мужской стерильности ms-3 и ms-4 в Cryptomeria japonica D. Don 2015 ; () –
Анализ случайных последовательностей бакклонов кедра японского, cryptomeria japonica 2015
Изменение скорости эволюции двух видов хвойных, Taxodium distichum (l.