Ель обыкновенная — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Рис. \(1\). Ель
У ели обыкновенной крона в виде конуса образована распростёртыми или поникающими ветвями. Кора у ели красновато-коричневого или серого цвета.
Хвоинки тёмно-зелёные, четырёхгранные, расположены одиночно, на дереве сохраняются \(5\)–\(7\) лет.
Рис. \(2\). Органы размножения ели
Ель обыкновенная начинает образовывать семена в возрасте \(25\)–\(30\) лет. На концах прошлогодних побегов образуются женские шишки. Они зеленоватые или фиолетово-красные, расположены вертикально. Зрелые шишки после высеивания семян опадают. Мужские шишки расположены ниже женских и имеют желтовато-бурую окраску.
Пыльца развивается в мужских шишках, семязачатки — в женских шишках. Ели цветут в мае, и шишки созревают в октябре того же года. Семена у ели крылатые, высыпаются из шишек весной (в марте-апреле).
Семена у ели крылатые, высыпаются из шишек весной (в марте-апреле).
Рис. \(3\). В лесу после сильного ветра
Ель имеет поверхностную корневую систему, которая расположена в верхнем слое почвы. Если ельник не защищён с подветренной стороны рощей или отдельно стоящими деревьями, то ветер часто валит ели. По сравнению с соснами ели растут в местах с более богатой питательными веществами почвой, но плохо переносят заболачивание почвы. Ель обыкновенная является очень теневыносливым растением. Может долго расти в тени других деревьев.
Продолжительность жизни ели обыкновенной составляет \(200\)–\(300\) лет. Она может достигать \(40\) м в высоту.
Древесину ели используют в производстве музыкальных инструментов, она является также ценным видом топлива.
Источники:
Рис. 1. Ель
Рис. 2. Органы размножения ели https://image.shutterstock.com/image-vector/-600w-1518657737.jpg
Рис. 3. В лесу после сильного ветра
|
Хвойные растения являются самыми древними из существующих растений на нашей планете. ХвояХвоя внешне походит на узкие игольчатые листья. Характерно наличие плотной кожицы, которую покрывает восковидная субстанция. Это необходимо для уменьшения испарения влаги голосеменными. К примеру, у ели хвоинки четырехгранные, но зачастую грани практически незаметны, и хвоинки выглядят уплощенными. Рисунок. Поперечный разрез хвои сосны обыкновенной Если разрезать хвоинку, она имеет форму неправильного ромба, причем самый плоский угол направлен книзу. Здесь расположена срединная жилка листа. Вдоль других граней иголочки заметны полосы белого цвета, образованные устьицами – дыхательными отверстиями, через которые осуществляется дыхание растений. Важным отличием хвойных пород от лиственных заключается в том, что у них черешок прочно связан с веткой и остается на ней, даже после отмирания иголки. Иголки отпадают через 6-7 лет. Они хорошо защищены от воздействия неблагоприятных факторов внешней среды толстым слоем воскового налета – кутикулой. Причем у многих видов налет настолько толстый, что хвоинки приобретают голубой оттенок. У хвойных нет истинных плодов и цветов. Они принадлежат к отделу голосеменных. Их семена прикреплены непосредственно к семенным чешуям, а те из них, которые собраны в женские шишки снабжены особыми крылышками. Покидая шишку, они планируют на крыльях, при вращении напоминая маленькие вертолетики. Хвойные шишкиВнешний вид шишек хвойных разнообразен и специфичен. Они могут отличаться по длине, форме, размещению в пространстве, цвету, структуре и форме спорофиллов, способу распространения семян, т. д. Но принципиальная структура шишек одинакова. Все шишки в основании имеют ось, которая обособлена от вегетативной части дерева и является коротким побегом с расположенными на нем спороносными листьями – спорофиллами. Различают женские и мужские шишки. Подавляющее большинство хвойных относятся к однодомным. У них и женские, и мужские шишки развиваются на одном растении. В большинстве случаев мужские шишки группами сосредоточены в пазухах листьев, иногда – на верхушках боковых побегов. Шишки женского пола отличаются компактным расположением, изредка они расположены поодиночке. Похожие материалы: |
 Их возраст исчисляется сотнями миллионов лет. Эволюция практически не повлияла на анатомическое строение хвои и шишек. При сравнении листьев хвойных, которые в простонародье называют иголками, с листьями цветковых растений, можно заметить, что при относительном однообразии хвоинок они имеют различную форму, размеры, окраску, а у некоторых видов совсем не выглядят, как привычная хвоя.
Их возраст исчисляется сотнями миллионов лет. Эволюция практически не повлияла на анатомическое строение хвои и шишек. При сравнении листьев хвойных, которые в простонародье называют иголками, с листьями цветковых растений, можно заметить, что при относительном однообразии хвоинок они имеют различную форму, размеры, окраску, а у некоторых видов совсем не выглядят, как привычная хвоя. Также устьица служат для испарения влаги, которую дерево впитывает из грунта даже в лютую стужу. Это объясняет тот факт, что ели, как и другие хвойные, нельзя пересаживать в осеннее время, так как корни не могут крепко укорениться, и вода практически не поднимается по стеблю к хвое, хотя дыхание происходит в том же режиме.
Также устьица служат для испарения влаги, которую дерево впитывает из грунта даже в лютую стужу. Это объясняет тот факт, что ели, как и другие хвойные, нельзя пересаживать в осеннее время, так как корни не могут крепко укорениться, и вода практически не поднимается по стеблю к хвое, хотя дыхание происходит в том же режиме.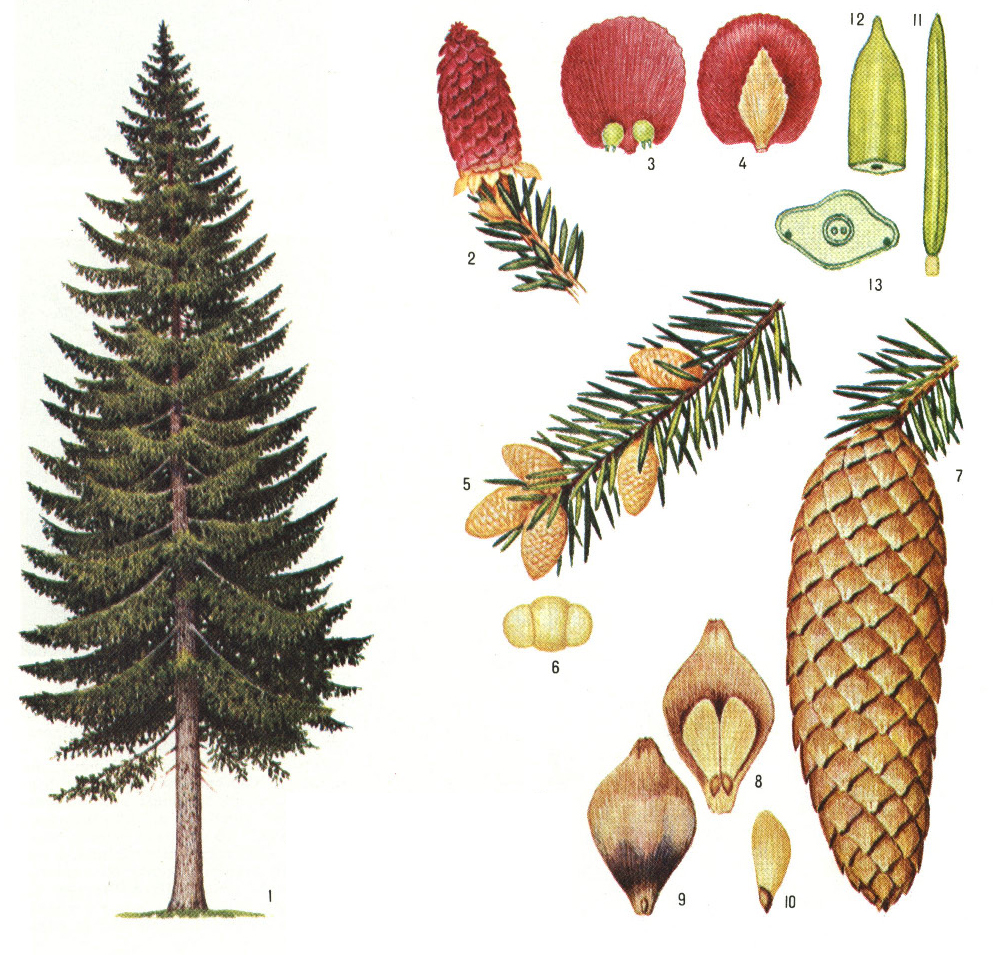 Это помогает им перемещаться подальше от материнского растения.
Это помогает им перемещаться подальше от материнского растения.Описать внешний вид сосна
Хвойное дерево сосна обыкновенная
Похожие статьиВ медицинских целях у можжевельника используют в основном плоды, то есть шишкоягоды. Собирать их лучше всего в период, когда они максимально созревают, это конец августа – начало сентября. Надо заметить, что у всех хвойных, в том числе у можжевельника, плодоношение по годам не одинаково и большое количество плодов образуется обычно раз в 3 – 4 года. Если вдруг вы пошли в лес и обнаружили можжевельник, густо усыпанный шишкоягодами, то соберите заранее и впрок, потому что на следующий год их может там не оказаться вообще. Сроки хранения этого сырья составляют 2 – 3 года.
Собирать их лучше всего в период, когда они максимально созревают, это конец августа – начало сентября. Надо заметить, что у всех хвойных, в том числе у можжевельника, плодоношение по годам не одинаково и большое количество плодов образуется обычно раз в 3 – 4 года. Если вдруг вы пошли в лес и обнаружили можжевельник, густо усыпанный шишкоягодами, то соберите заранее и впрок, потому что на следующий год их может там не оказаться вообще. Сроки хранения этого сырья составляют 2 – 3 года.
. В культуре, правда, встречаются и другие виды ели, например, голубые ели. В лесу можно встретить растение очень похожее на ель, которое называется пихта. Иголочки пихты более мягкие, на концах не такие острые, более округлые и плоские, чем у ели. У ели иголочки – это самое ценное медицинское сырье, гораздо более ценное, чем еловые шишки.Смолу сосны собирают с живых растений и называют “живица”. В промышленных масштабах с одного дерева можно собрать несколько килограммов смолы. Конечно же, после этого дерево погибает, поэтому делается это в тех местах, в тех участках леса, которые предназначены для рубки. Сосновую смолу издавна использовали вместо жевательной резинки. За счет того, что в ней содержится огромное количество витаминов, биологически активных веществ, эфирных масел ее использовали при борьбе с заболеваниями, связанными с недостатком витаминов, то есть при авитаминозах, а еще при воспалительных процессах ротовой полости и горла.
Конечно же, после этого дерево погибает, поэтому делается это в тех местах, в тех участках леса, которые предназначены для рубки. Сосновую смолу издавна использовали вместо жевательной резинки. За счет того, что в ней содержится огромное количество витаминов, биологически активных веществ, эфирных масел ее использовали при борьбе с заболеваниями, связанными с недостатком витаминов, то есть при авитаминозах, а еще при воспалительных процессах ротовой полости и горла.
Пигмеа. Маленькое чудо для альпийской горки, размер которого всего лишь 40 см в высоту. Многовершинное деревце с золотистыми иголками.Главное, не пропустить заболевания, связанные с грибковыми заболеваниями типа «Шютте обыкновенное». Бороться с ними можно с помощью применения фунгицида «Тилт». Признаком заболевания раком является засыхание хвои в мае и отмирание побегов в летний период. Регулярным многократным поливом рас вором фундазола можно попытаться спасти дерево. Часто среди хвои появляется белый пушок.
Поэтому во многих странах разрабатывались различные методики выращивания необходимого количество крепких молодых деревцев, способных хорошо расти в разных климатических условиях.Сосновая древесина широко применяется в производстве мебели. Используется она и в строении судов и вагонов. Из нее сегодня возводят портовые сооружения, дамбы и причалы. Сосновый лес даже называли «корабельной рощей» или «мачтовым лесом». А суда – «плавучими соснами». Сосновая смола интенсивно использовалась корабельщиками для обработки канатов, судов и лодок. Все это говорит о высоких характеристиках сосновой древесины.
Древесина обыкновенной сосны плотная, ядровая, содержащая много древесной смолы. Молодые посадки характеризуются прямослойной древесиной, которая с годами переходит в косослойную. Плотность древесины и ее механические свойства, которые важны в строительстве, зависят от ряда факторов, в частности: влажность грунта. Так, сосна, растущая на сухой почве, имеет более плотную и устойчивую к повреждениям древесину.
40 мБотаническое название:
О полезных бактерицидных свойствах сосны, ели и других хвойных растений было известно людям давным-давно. Например, ученые обнаружили, что в глиняных клинописных табличках древних шумеров, возраст которых датируется около 5000 лет, есть упоминание о полезных свойствах сосны. Древние египтяне использовали смолу, то есть живицу сосны и ели для приготовления бальзамирующих составов. Удивительно, прошло более 3000 лет, а эти составы не потеряли своих бактерицидных свойств. В медицине используются в основном молодые побеги ели. Лучше всего собирать хвою весной, в тот момент, когда иголочки еще мягкие и отличаться по цвету от старых – они более светлые и яркие. Хвоя ели так же, как и хвоя сосны, содержит очень большое количество полезных элементов. В первую очередь это эфирные масла, фитонциды, различные микроэлементы. Хвоя ели очень богата дубильными веществами, и, самое главное – здесь содержится большое количество витамина C, даже больше, чем в хвое сосны. Именно поэтому молодые побеги ели во время войны использовали как мощное средство против цинги. Их жевали, добавляли в салат, в различные другие блюда. Весной, прогуливаясь по лесу, если вы обнаружили на ели молодые хвоиночки, попробуйте их на вкус – они будут кисловатыми.
В медицине используются в основном молодые побеги ели. Лучше всего собирать хвою весной, в тот момент, когда иголочки еще мягкие и отличаться по цвету от старых – они более светлые и яркие. Хвоя ели так же, как и хвоя сосны, содержит очень большое количество полезных элементов. В первую очередь это эфирные масла, фитонциды, различные микроэлементы. Хвоя ели очень богата дубильными веществами, и, самое главное – здесь содержится большое количество витамина C, даже больше, чем в хвое сосны. Именно поэтому молодые побеги ели во время войны использовали как мощное средство против цинги. Их жевали, добавляли в салат, в различные другие блюда. Весной, прогуливаясь по лесу, если вы обнаружили на ели молодые хвоиночки, попробуйте их на вкус – они будут кисловатыми.
Смола сосны является еще и ценнейшим химическим сырьем. Из нее, например, получают скипидар, канифоль. Канифоль используют и в музыкальной промышленности для смазывания смычков, и при пайке различных металлических изделий. А вот окаменевшая смола – это янтарь.Сосна обыкновенная. Большое красивое дерево с фантастической аурой, создающей особенную атмосферу вокруг себя. Целебный аромат, идущий от сосны, делает приятным отдых в её тени.
А вот окаменевшая смола – это янтарь.Сосна обыкновенная. Большое красивое дерево с фантастической аурой, создающей особенную атмосферу вокруг себя. Целебный аромат, идущий от сосны, делает приятным отдых в её тени.
Описание сосны обыкновенной
Это вредное действие появившейся тли, называемой сосновый хермес. Борьбы с ней проводится опрыскиванием раствором ровикурта или актеллика. При желании можно освоить обрезку ветвей для формирования нужной кроны.
Одним из инновационных методов является способ выращивания саженцев сосны с закрытой корневой системой. Хотя в России до сих пор практикуется выращивание саженцев методом открытой корневой системы в незащищенном грунте.
Однако посадки сосен применяются и в других целях. Так, своеобразная сосновая корневая система позволяет предупредить эрозию почвы, обеспечивает оптимальный уровень влажности, предохраняет обрывы и овраги от обсыпания.
Хвойное растение сосна
Данный представитель хвойных хорошо размножается семенами. Для этого необходим хороший грунт и обилие солнца. Лучше всего пересаживать саженцы в возрасте 3-7 лет.
Для этого необходим хороший грунт и обилие солнца. Лучше всего пересаживать саженцы в возрасте 3-7 лет.
Сосна обыкновенная образует целый ряд форм, которые различаются между собой строением кроны, цветом и формой шишек. Встречаются разновидности с плакучей и пирамидальной кроной. Окраска иголок у молодых побегов может быть золотистая, беловатая или серебристая. Кора встречается чешуйчатая или пластинчатая.
Характеристика сосны обыкновенной
Средняя продолжительность жизни:
сосна обыкновенная (Pinus silvestris)
Мужские шишки сосны желтого цвета (т. к. в них много желтой пыльцы) , располагаются у основания молодых веточек.
Отвары и настои еловой хвои используют при таких заболеваниях, как ангины, тонзиллиты для полоскания и приема внутрь. Чаще всего, в отличие от сосны, ель используют при подагре, ревматизме, различных заболеваниях, связанных с образованием камней в почках и мочевыводящей системе.
Препараты хвои сосны обладают целым рядом полезных медицинских свойств. Их используют как мочегонное, желчегонное, общеукрепляющее, кровоостанавливающее средство. Кроме того, они обладают еще и дезинфицирующими и отхаркивающими свойствами. Отвар сосновой хвои используют при бронхитах, заболеваниях верхних дыхательных путей, астме. Их пьют или используют в качестве ингаляций.
Их используют как мочегонное, желчегонное, общеукрепляющее, кровоостанавливающее средство. Кроме того, они обладают еще и дезинфицирующими и отхаркивающими свойствами. Отвар сосновой хвои используют при бронхитах, заболеваниях верхних дыхательных путей, астме. Их пьют или используют в качестве ингаляций.
Может быть, сосна, с любовью и заботой посаженная на дачном участке, когда-нибудь станет одним из самых древних деревьев планеты, напоминая знаменитого Мафусаила, растущего на протяжении почти пяти тысячелетий в национальном парке Калифорнии.
Чем отличается дерево сосна?
Все этапы выращивания сосны можно повторить в домашних условиях самостоятельно, используя плёнку и специально организованную школку для подращивания сеянцев. В конце очень важно не забыть пройти стадию закаливания саженцев перед высадкой их в открытый грунт. Этот процесс не очень быстрый. В лесных хозяйствах практикуется использовать схему, по которой на протяжении двух лет сеянец растёт в семенном отделении и еще столько же в школке.
Необходимость освоения дорогостоящего современного метода связана с получением растений с крепкой корневой системой. Именно от её величины в сравнении с зеленой частью зависит процент выживаемости после посадки на постоянное место. Часто обычные дачники идут в лес и копают дерево, которое погибает на второй год жизни. Чтобы избежать этого, необходимо строго выполнять все этапы технологии выращивания сеянцев, а именно:
В качестве декоративной культуры используется крайне редко. Чаще применяется в озеленении загородных усадеб, профилакториев, санаториев. Высаживается в смешанных насаждениях, одиночно, массивами или группами. Могут использоваться в озеленении лесопарков и садово-парковых территорий, для обсадки загородной дороги, особенно на бедной песчаной почве. В культуре произрастает группой или одиночно в крупных парках, садах и скверах. Декоративность молодой посадки маловыразительная. Высокую декоративность деревья приобретают к ста годам, когда ствол в своей верхней части начинает покрываться тонкой корой оранжевого цвета, которая придает нарядность и привлекательность дереву. Эстетические характеристики отдельных сосен усиливаются по мере того, как изреживается древостой. Особенной неповторимостью обладают старые сосны, притягивающие взгляды издалека. В условиях отсутствия загрязнения воздуха сосна обыкновенная обладает высокими санитарно-гигиеническими характеристиками.
Эстетические характеристики отдельных сосен усиливаются по мере того, как изреживается древостой. Особенной неповторимостью обладают старые сосны, притягивающие взгляды издалека. В условиях отсутствия загрязнения воздуха сосна обыкновенная обладает высокими санитарно-гигиеническими характеристиками.
Сосна плохо реагирует на загрязненный городской воздух, хотя и произрастает там довольно часто. За 2 года жизни в городе смолистая поверхность иголок покрывается пылью и копотью, что мешает фотосинтезу растения.
Ареал сосны обыкновенной
Растение сосна имеет широкий ареал, который простирается на разных, с экологической точки зрения, районах, поэтому вид характеризуется большим количеством экотипов. На сегодняшний день экологами выделяется более 30 таких экотипов. Например, ангарская сосна, произрастающая в бассейне р. Ангара, является экотипом сосны обыкновенной. Изучение и наблюдение за выращенными в одинаковых условиях саженцев сосны обыкновенной разного происхождения показывает различие растений в способностях к засухо- и холодоустойчивости, к росту, сопротивляемости заболеваниям. Также эти растения могут отличаться морфологическими признаками, такими как: форма кроны, длина иголок, строение ствола и пр. Однако все эти признаки склонны к изменениям и не используются для выделения вида.
Также эти растения могут отличаться морфологическими признаками, такими как: форма кроны, длина иголок, строение ствола и пр. Однако все эти признаки склонны к изменениям и не используются для выделения вида.
200 лет
Сосна обыкновенная: применение
Родина:
У папоротников споры находятся в мешочках – спорангиях, располагающихся на нижней стороне листа.
Но у препаратов ели есть и свои противопоказания. Например, их не рекомендуется употреблять в большом количестве людям, которые страдают некоторыми заболеваниями почек. В основном это нефриты, нефрозы и заболевания близкие к этим.
Молодые почки сосны собирают обычно весной в тот момент, когда хвоиночки еще не до конца вылезли из своих почечных чешуй. Их срезают ножницами, сушат, и в дальнейшем из них готовят настои и отвары. А вот хвою сосны можно собирать практически в любой период, даже зимой. Собирать хвою нужно с тоненьких молодых веточек, диаметр которых составляет не больше 2 – 3 миллиметров. Такая хвоя хранится не долго, обычно 2 – 3 месяца; из нее можно готовить отвары, настои. Ее мелко режут и заливают холодной водой. В этом случае настой готовится в течение 12 часов, и пьется по полстакана 2 – 3 раза в день.
Такая хвоя хранится не долго, обычно 2 – 3 месяца; из нее можно готовить отвары, настои. Ее мелко режут и заливают холодной водой. В этом случае настой готовится в течение 12 часов, и пьется по полстакана 2 – 3 раза в день.
Видео-инструкция по размножению хвойных деревьев черенком:
Посадка на постоянное место производится в заранее подготовленное место, желательно на солнце, которое очень любит сосна. В ямку вносятся минеральные удобрения, производится полив и мульчирование торфом.
Фотогалерея: сосна обыкновенная (нажмите на картинку для увеличения):
udec.ru
Как провести размножение сосны в домашних условиях?
Подготовку семян перед посадкой. С этой целью используют бережное удаление крылышек за счет вращения в воде. Для активизации процессов пробуждения и роста семян их помещают в холодильную камеру со строго поддерживаемой температурой в -1°и влажностью. В таком состоянии они могут храниться несколько лет, не теряя своей всхожести. В домашних условиях очень часто применяется метод снегования семян. Во второй половине марта холщовый мешок с семенами помещается в снег, закрытый опилками, а потом ещё одним слоем снега.
Во второй половине марта холщовый мешок с семенами помещается в снег, закрытый опилками, а потом ещё одним слоем снега.
В настоящее время почти на каждом участке растёт какое-нибудь хвойное дерево. Ими украшены городские скверы, парки, улицы. В любую пору года они привлекают к себе взоры прохожих. И если близлежащий лес состоит в основном из посадок хорошо известной сосны обыкновенной, то на ухоженных частных владениях, территориях ботанических садов и парков можно наблюдать за оригинальными деревцами, относящимися к определённым сортам сосны, число, которых, составляет около 100 единиц. Конечно, просто так они не растут.
Внешний вид сосны
Быстрорастущие хвойные растения, сосна обыкновенная в том числе, без применения обрезки опережают окружающие деревья и успешно доминируют над ними в росте. Обрезка хвойных выполняется для формирования и поддержания структуры дерева и увеличения продолжительности его жизни. Грамотная обрезка снижает вероятность возникновения дефектов, структурных отклонений дерева. Кроме того, сформированная крона предотвращает падение сосны в результате негативного воздействия погодных условий. Сломанные, засохшие или с проявлением болезней ветви немедленно удаляются, что предотвращает распространение грибковых заболеваний. Также может быть удалена живая ветвь. Это бывает в исключительных случаях и необходимо для обеспечения попадания солнечных лучей и циркуляции воздуха внутри кроны.
Кроме того, сформированная крона предотвращает падение сосны в результате негативного воздействия погодных условий. Сломанные, засохшие или с проявлением болезней ветви немедленно удаляются, что предотвращает распространение грибковых заболеваний. Также может быть удалена живая ветвь. Это бывает в исключительных случаях и необходимо для обеспечения попадания солнечных лучей и циркуляции воздуха внутри кроны.
Произрастая в неблагоприятных условиях, например на болоте, сосна обыкновенная может остаться карликом. При этом даже столетние экземпляры могут не превышать 1 м в высоту. Сосна – светолюбивое растение, морозо- и жароустойчивое. Из всех представителей древесных пород, растущих на песчаных почвах, сосна обыкновенная наиболее устойчива к нехватке влаги. В таких условиях корни способны проникнуть в почву на глубину 6 м. Поэтому даже в условиях засухи могут снабжать дерево водой. Данная способность растений и обусловило разную корневую систему разных популяций. На засушливых территориях у дерева хорошо развивается стержневой корень, а в условиях близкого залегания грунтовых вод, корневая система формируется, в основном, боковыми корнями, разветвляющимися во все стороны.
Размножение:
Размножение сосны своими руками
Сибирь, Урал, Европа
school-collection.edu.ru/files/materials/640/totals_biology.xls
Несмотря на то, что сосна и ель являются близкими родственниками, они даже относятся к одному семейству – семейству сосновых, леса из них очень разнообразны по своему внешнему виду. Например, сосновый лес светлый, прозрачный и как бы наполнен воздухом. Именно поэтому сосновые леса, их еще по-другому называют – сосновый бор, очень часто вдохновляли художников на написание этих пейзажей. А лес еловый, наоборот, темный и мрачный. Это связано с тем, что ель образует очень густую крону, которая практически не пропускает солнечного света. Именно поэтому в еловых лесах кроме самой ели ничего не растет.
В медицинских целях у сосны используют еще и пыльцу. Пыльца у сосны созревает в мужских шишках. Эти шишки, в отличие от женских, расположены обычно на нижних побегах и собирать их достаточно легко. Время, оптимальное для сбора, тоже обнаружить очень просто. Если в конце мая – начале июня подойти к сосне и ударить по нижним веткам, то в период созревания пыльцы над этими ветками поднимется целое облако желтой пыли – это и есть пыльца. Ее собирают, срезая ножничками или ножом, отделяют хвоинки, сушат и используют так же в виде настоев и отваров. Пыльца сосны – это часто используемая и встречаемая биологически активная добавка. Она является в первую очередь общеукрепляющим и иммуностимулирующим свойством. Кроме того, считается, что препараты пыльцы сосны обладают омолаживающим свойством и способствуют увеличению подвижности суставов.
Если в конце мая – начале июня подойти к сосне и ударить по нижним веткам, то в период созревания пыльцы над этими ветками поднимется целое облако желтой пыли – это и есть пыльца. Ее собирают, срезая ножничками или ножом, отделяют хвоинки, сушат и используют так же в виде настоев и отваров. Пыльца сосны – это часто используемая и встречаемая биологически активная добавка. Она является в первую очередь общеукрепляющим и иммуностимулирующим свойством. Кроме того, считается, что препараты пыльцы сосны обладают омолаживающим свойством и способствуют увеличению подвижности суставов.
- Хвойные – деревья и кустарники, листья которых видоизменены в иглы или чешуи. Семена созревают внутри деревянистых шишек или сочных шишкоягод. Все представители содержат большое количество эфирных масел и смолу. Многие виды имеют медицинское значение.
- Кроме семенного размножения сосен при желании можно овладеть более сложными методами прививки.
Посев семян на линиях автоматического посева. Специальные ячейки заполняются торфом, он уплотняется и в создаваемые имеющимся устройством лунки, высеваются по два семечка. Они закрываются тонким слоем опилок и помещаются в теплицу. Всходы появляются на двадцатый день.
Специальные ячейки заполняются торфом, он уплотняется и в создаваемые имеющимся устройством лунки, высеваются по два семечка. Они закрываются тонким слоем опилок и помещаются в теплицу. Всходы появляются на двадцатый день.
Чтобы вырастить красивое деревце надо воспользоваться предложениями специализированных фирм, продающих саженцы или вырастить его самостоятельно. В любом случае придётся обратиться за полезной информацией по уходу за ним.
Данный вид хвойных широко распространен в Сибири и Европе. Образует сосновые леса на песчаной или супесчаной почве, может встретиться на торфянистом и очень редко на глинистом грунте. Это широко распространенное дерево Евразии. Его можно встретить от Испании и Великобритании на восток до р. Алдан и р. Амура в Сибири. На севере дерево произрастает до Лапландии, а на юге встречается в Китае и Монголии. Образует, как чистые насаждения, так и вместе с другими хвойными, дубом, березой и осиной. Растение нетребовательно к почвенно-грунтовым условиям и очень часто растет на непригодных для других видов участках: песках, болотах.
Средняя продолжительность жизни сосен составляет порядка 200 лет. Отдельные экземпляры при благоприятных условиях живут до 400 лет.
семенами, прививкой
Освещение:
попробуй тут
Но есть еще другое семейство, которое также относится к хвойным, это семейство кипарисовые. И помимо южных представителей этого семейства, таких как туя и кипарис, которые в нашей средней полосе не встречаются, есть одно дерево, или кустарник, который также используется в медицине – это можжевельник.
Из пыльцевых шишек сосны никогда не получатся шишки настоящие, потому что это мужские шишки. А вот женские шишки находятся на других ветках и обычно эти ветки расположены более высоко. Надо сказать, что шишка сосны и шишка ели очень сильно друг от друга отличаются. Шишка сосны покороче и чешуйки, которые прикрывают семена, более твердые, жесткие и деревянистые. Женские шишки сосны в медицине не используются. Шишки ели, наоборот, вытянуты и в свежем (не в сухом) виде они имеют вид морковки. Чешуйки у них более тонкие и они плотно примыкают к осевой части, закрывая семена, которые там спрятаны.
Чешуйки у них более тонкие и они плотно примыкают к осевой части, закрывая семена, которые там спрятаны.
Сорта сосен для ландшафтного дизайна
Медицинские свойства всех хвойных растений в первую очередь обусловлены теми веществами, которые входят в их состав. В основном это эфирные масла. Именно поэтому хвойные растения так любят банщики и приверженцы ароматерапии. Самое распространенное и самое заметное из всех хвойных, которые у нас растут, это, наверное, сосна. Сосна – дерево высотой до 40 метров, к этому роду относится большое количество видов растений. Самый распространенный вид – это
Научные исследования показали, что сосны относятся к реликтовым видам деревьев, пришедших к нам из эпохи динозавров. Но их первобытная красота дошла к нам практически без изменений вместе с многочисленными красивыми легендами о ней. Существует большое количество видов этого дерева. Они имеют разную высоту, форму кроны, размер иголок.
- После появления всходов растения подкармливаются, например, удобрением Кемира-универсал 2, содержащим набор микроэлементов.
 Регулярно поливаются и только через 6 месяцев подросшие саженцы помещают в другое более прохладное помещение для закаливания.
Регулярно поливаются и только через 6 месяцев подросшие саженцы помещают в другое более прохладное помещение для закаливания. - Вечнозелёное хвойное дерево, цветущее в мае, относится к семейству сосновых растений Pinaceae. Оно включает в себя ели, пихты, лиственницы, кедры, и имеет латинское название Pinus sylvestris L. Высота сосны может достигать 50 м, а продолжительность жизни составляет 500 и даже более лет. Листья представляют собой игольчатые хвоинки длиной 5,7 см. Они тёмно-зелёного цвета, выпуклые в верхней части и имеющие желобок на внутренней стороне.
- Сибирский ареал занимает площадь около 5,7 млн. км2. Наиболее крупные сосновые массивы сосредоточены в бассейне р. Ангары, в верховьях Иртыша, Оби, Подкаменной Тунгуски. В северной части ареала распространение сосны поднимается на уровень 1000 м над уровнем моря, а на юге – до 1500 м над уровнем моря.
- Растут они быстро, особенно существенный прирост с года до 100 лет – 50-70 см. По этому показателю данный представитель хвойных уступает только лиственнице. Плодоносить начинает с 15-летнего возраста. В условиях густой посадки – с 40 лет. Обильные урожаи, как правило, могут повторяться через 4-7 лет.
 Регулярно поливаются и только через 6 месяцев подросшие саженцы помещают в другое более прохладное помещение для закаливания.
Регулярно поливаются и только через 6 месяцев подросшие саженцы помещают в другое более прохладное помещение для закаливания. Плодоносить начинает с 15-летнего возраста. В условиях густой посадки – с 40 лет. Обильные урожаи, как правило, могут повторяться через 4-7 лет.
Плодоносить начинает с 15-летнего возраста. В условиях густой посадки – с 40 лет. Обильные урожаи, как правило, могут повторяться через 4-7 лет.Синоним –
светолюбивая
OgorodSadovod.com
Хвойные растения. Полезные свойства сосны, ели и можжевельника.
Детальное описание иллюстраций:Можжевельник обыкновенныйМолодые с недозревшими семенами шишки ели используют в медицине. Из них готовят отвары, которые используют при различных заболеваниях, связанных с воспалительными процессами рта, горла, дыхательных путей – это тонзиллиты, ларингиты, бронхиты, ангины. Их используют в виде полосканий и применяют также и наружно. В отличие от сосны, которая используется и в официальной, и в народной медицине, ель используется только в медицине народной, но там она занимает свое достойное место.Сосна обыкновенная
Хочется посадить их как можно больше, но здравый смысл и опыт соседей, всегда подскажет какой сорт выбрать.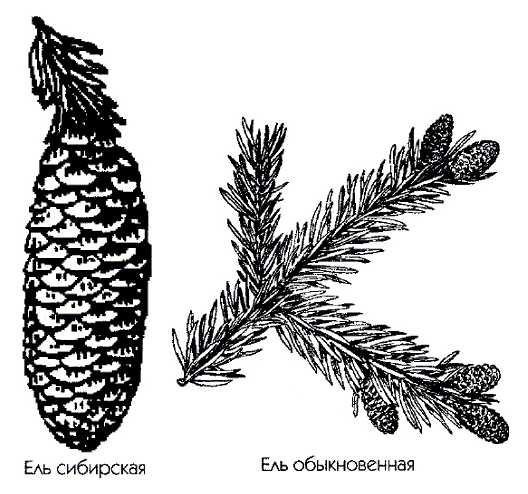 Наиболее популярными декоративными видами сосны на небольших дачных участках являются следующие:
Наиболее популярными декоративными видами сосны на небольших дачных участках являются следующие:
Стандартной высоты, необходимой для посадки саженца на постоянное место, он достигает при таком способе выращивания через год. Она составляет примерно 12 см при очень крепкой корневой системе. В открытом грунте такой размер достигается только через три года.
Крепятся иголки по две и растут до 5 лет. Почки сосны выглядят в виде молодых побегов, расположенных на верхушках стволов. Имеют розовый цвет с буроватым оттенком. Зелёные шишки первого года, почки, побеги, хвоя, живица, пыльца обладают лечебными свойствами, и многие века используются в рецептах медицины разных народов.
Ветви и ствол сосны пронизан смоляными ходами, которые наполнены смолой, обычно называемую «живицей». «Живица» имеет большое значение для растения: заживляет нанесенные раны, отпугивает вредителей. Добывают такую живицу путем подсочки. Используется она для получения канифоли, скипидара. Главное, что получить ее можно не только с живого дерева, но и с соснового пня. Воздух в сосновом бору («смолистый») богат озоном и не содержит микробов. Сосновые леса издавна славятся благоприятными для человека свойствами.
Воздух в сосновом бору («смолистый») богат озоном и не содержит микробов. Сосновые леса издавна славятся благоприятными для человека свойствами.
Хорошо растет на супесчаных и песчаных почвах. Крайне редко встречается в степных южных районах. Поэтому в последнее время сосну обыкновенную часто высаживают в лесозащитных полосах по склонам оврагов, на песках, в степных балках.
сосна лесная
Почва:
Ель обыкновенная. Весной на концах ветвей ели обыкновенной можно увидеть ярко-красные, величиной с наперсток женские шишечки (1). Тонкие, нежные чешуйки таких шишек похожи на лепестки цветков. На внутренней поверхности лепестков появляются две семяпочки, из которых впоследствии вырастают крохотные, снабженные пленчатыми крылышками семена (2). Мужские шишечки ели обыкновенной мельче женских (3). На наружной стороне их чешуек хорошо заметны пыльники (4).– вечнозеленый кустарник семейства кипарисовые. Растет в подлеске хвойных и хвойно-мелколиственных лесов. Семена расположены в сочных мясистых шишкоягодах, которые используются в медицине, а также для приготовления джина. Все части растения, включая древесину, содержат большое количество фитонцидов.На территории нашей страны в диком виде растет в основном один вид – это. Хвоинки сосны обыкновенной собраны по две и не очень длинные. Есть сосны, у которых хвоинки достигают 15 – 20 см. Еще одно растение, которое тоже многим знакомо, в простонародии его называют “кедр”, хотя на самом деле это Сосна сибирская. Иголочки сосны сибирской собраны в пучок по пять штук. Семена кедра использовались в медицине давным-давно. В Сибири хорошо известно, что кедровые орешки содержат большое количество белка, витаминов и различных микроэлементов. Именно поэтому их использовали не просто в пищу, а очень часто измельченные и с небольшим добавлением воды они использовались в качестве заменителя грудного молока при недостатке его у матери.
Все части растения, включая древесину, содержат большое количество фитонцидов.На территории нашей страны в диком виде растет в основном один вид – это. Хвоинки сосны обыкновенной собраны по две и не очень длинные. Есть сосны, у которых хвоинки достигают 15 – 20 см. Еще одно растение, которое тоже многим знакомо, в простонародии его называют “кедр”, хотя на самом деле это Сосна сибирская. Иголочки сосны сибирской собраны в пучок по пять штук. Семена кедра использовались в медицине давным-давно. В Сибири хорошо известно, что кедровые орешки содержат большое количество белка, витаминов и различных микроэлементов. Именно поэтому их использовали не просто в пищу, а очень часто измельченные и с небольшим добавлением воды они использовались в качестве заменителя грудного молока при недостатке его у матери.
Глаука. Сосна имеет красивые пушистые ветви при ширине конической кроны в 1 м и высотой до 2 м. Цвет хвои серо-голубой.
Поэтапное выращивание саженцев методом закрытой корневой системы полностью исключает её травмирование и обеспечивает 100% приживаемость. Причем саженцы, имеющие закрытую корневую систему можно пересаживать не только весной или осенью, но и в другие сроки, когда рост надземной части деревца только начался или ещё не прекратился.
Причем саженцы, имеющие закрытую корневую систему можно пересаживать не только весной или осенью, но и в другие сроки, когда рост надземной части деревца только начался или ещё не прекратился.
Шишки сосны имеют яйцевидную форму и состоят из чешуек, закрывающих семена. Только на второй сезон они раскрываются, и семена можно использовать для новых посадок. Чаще всего семена имеют одно крыло, помогающее ему перемещаться в воздушных потоках. У некоторых видов сосен крылышки у семян отсутствуют, и оно имеет деревянную оболочку в виде твёрдого орешка, например, плоды кедровой сосны, хорошо всем известной.
В медицине широко применяются почки, которые необходимо успеть собрать ранней весной, пока они не распустились. Почки содержат эфирные масла, смолы, крахмал, дубильные и горькие вещества. Сосновая хвоя содержит в большом количестве каротин и витамин С. Благодаря ценности древесины, сосновые леса считаются главным объектом лесоэксплуатации.
Очень часто можно встретить информацию, что сосна – двудомное растение. Это ошибка, на самом деле, это однодомное растение, то есть с преобладанием цветков или мужского, или женского пола. Таким образом, на одном дереве, преимущественно содержатся женские цветки, а на другом – преобладают мужские соцветия. Женские цветки располагаются на концах побегов и имеют форму небольшой шишечки. А мужские находятся возле основания побега. Считается, что преобладание соцветий определенного пола – наследственный фактор. Но, оказывается, в зависимости от условий, в которых произрастает дерево, его «пол» может поменяться.
Это ошибка, на самом деле, это однодомное растение, то есть с преобладанием цветков или мужского, или женского пола. Таким образом, на одном дереве, преимущественно содержатся женские цветки, а на другом – преобладают мужские соцветия. Женские цветки располагаются на концах побегов и имеют форму небольшой шишечки. А мужские находятся возле основания побега. Считается, что преобладание соцветий определенного пола – наследственный фактор. Но, оказывается, в зависимости от условий, в которых произрастает дерево, его «пол» может поменяться.
Дерево сосна является одной из ценнейших пород хвойных в нашей стране. Достигая 35-40 м в высоту, относится к деревьям первой величины. Окружность ствола достигает 1 м. Покрыт он красновато-бурой, с бороздками, отслаивающейся корой. У основания ствола кора намного толще той, которая расположена вверху. Такая «задумка» природы несет в себе защитную функцию, защищая дерево от перегрева и низового пожара. У сосен, которые растут в сомкнутых древостоях, ствол более стройный с ажурной кроной.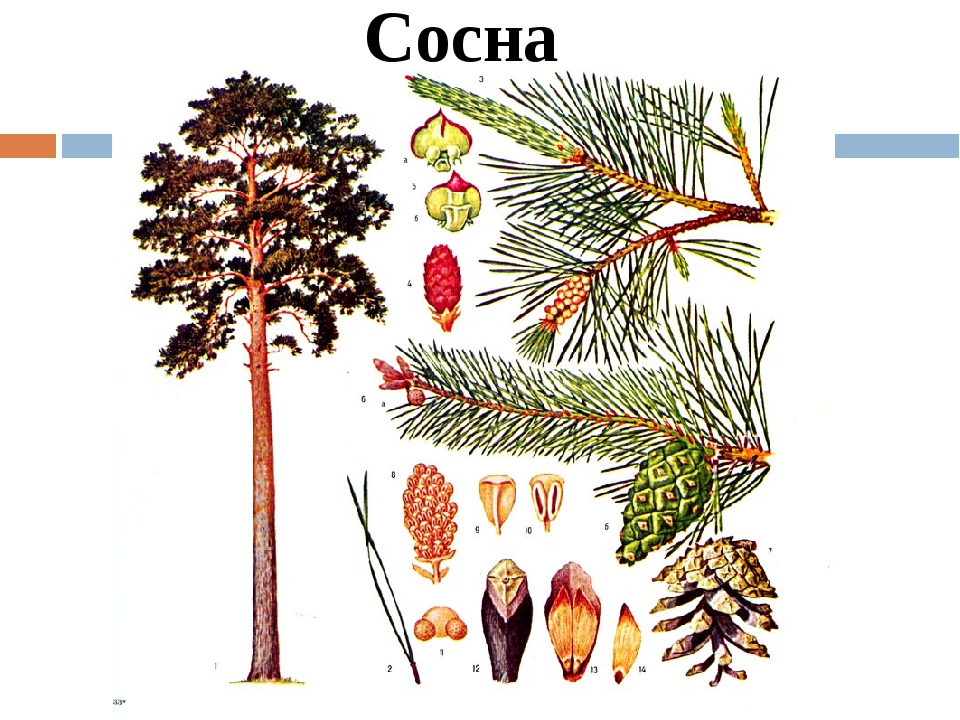 Пока дерево молодое, крона имеет конусовидную форму. С возрастом она круглеет, становится более шире, а в старости приобретает плоскую или зонтиковидную форму. Сосновая хвоя имеет сизо-зеленый цвет. Она достаточно плотная, часто торчащая, изогнутая, собранная в пучки по 2 иголки. Длина 4-7 см. Хвоинки заостренные, чуть сплюснутые, имеют тонкую продольную полосу. Живет хвоя 3 года. Осенью, чаще в сентябре, часть хвои опадает. Перед этим хвоинки приобретают желтый цвет, от чего крона выглядит пестрой.песчаная, супесчаная
Пока дерево молодое, крона имеет конусовидную форму. С возрастом она круглеет, становится более шире, а в старости приобретает плоскую или зонтиковидную форму. Сосновая хвоя имеет сизо-зеленый цвет. Она достаточно плотная, часто торчащая, изогнутая, собранная в пучки по 2 иголки. Длина 4-7 см. Хвоинки заостренные, чуть сплюснутые, имеют тонкую продольную полосу. Живет хвоя 3 года. Осенью, чаще в сентябре, часть хвои опадает. Перед этим хвоинки приобретают желтый цвет, от чего крона выглядит пестрой.песчаная, супесчаная
В отличие от ели и других хвойных растений можжевельник обычно не вырастает больше 3 – 5 метров. Самое большое можжевеловое дерево, которое описано ботаниками, расположено в Норвегии и высота его 17 метров; этому дереву около ста лет. В наших лесах обычно высота можжевельника 1,5 – 3 метра. Но, если вы посадите его у себя на участке, он может оказаться и более высоким. Внешний вид можжевельника очень сильно отличается от внешнего вида ели. Во-первых, у него совершенно другие хвоинки – они более короткие и значительно более острые, чем хвоинки ели. Кроме того, веточки можжевельника короткие и расположены ближе к основному стволу. Очень часто можжевельник представлен не одним стволом, а сразу двумя – тремя, которые отходят от одного корня.
Во-первых, у него совершенно другие хвоинки – они более короткие и значительно более острые, чем хвоинки ели. Кроме того, веточки можжевельника короткие и расположены ближе к основному стволу. Очень часто можжевельник представлен не одним стволом, а сразу двумя – тремя, которые отходят от одного корня.
Ель обыкновенная
Семена сосны обыкновенной значительно меньше по размеру и в пищу не используются, во всяком случае, человеком. Зато в медицинских целях используются многие другие части этого растения. Это и хвоя, и молодые почки, и пыльца и, самое главное – смола.
indasad.ru
Помогите пожалуйста! Мне нужно описать внешний вид мужских шишек и спороносящего папоротника!
Штирлиц
Нана. Голубая сосна, высотой до 1 м с многовершинной кроной. Сорт отличается красивыми пурпурными шишками.
Уход за молодыми посевами не составляет большого труда. Регулярный полив им не требуется.
наташа (левенцова)сафонова
В настоящее время требования коммерческих видов деятельности приводят к постоянно растущему увеличению промышленного выращивания саженцев сосны.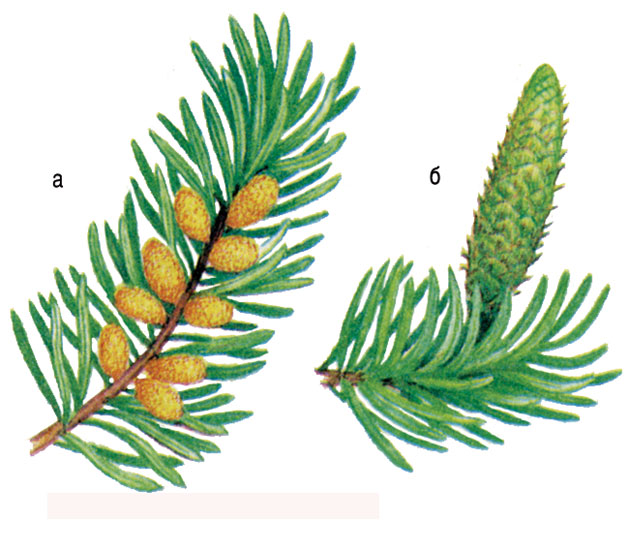 Восстановление лесных плантаций, потребность ландшафтных дизайнеров удовлетворить частными предложениями невозможно.
Восстановление лесных плантаций, потребность ландшафтных дизайнеров удовлетворить частными предложениями невозможно.
Сосна обыкновенная относится к самым древним лекарственным растениям. Ее хвоя входила в состав припарок и компрессов еще 5000 лет назад. В Древнем Египте сосновая смола находилась в бальзамирующих составах. Кстати, даже сейчас, спустя 3000 лет, эти составы не потеряли свои бактерицидные свойства. В Риме и Греции хвою использовали для лечения простуд. А на Руси для дезинфекции ротовой полости, укрепления зубов и десен было принято жевать сосновую смолу.
внешний вид пыльцы шишки рисунок и описание
…зовут меня Вовка…
Цветет сосна обыкновенная в конце мая, когда температура воздуха уже высокая. Опыление происходит за счет ветра. Само оплодотворение наступит только на следующий год. В период опыления на деревьях можно увидеть желтый налет. Это и есть сосновая пыльца. По большому счету, растение сосна отличается хорошей опыляемостью.
 Такое возможно за счет воздушных мешочков, которые имеет сосновая пыльца, при помощи которых она и разносится ветром на большие расстояния. Срок опыления изменяется в зависимости от погоды. При ясной солнечной погоде пыльца может разлететься за 3-4 дня. В дождь этот процесс затягивается.
Такое возможно за счет воздушных мешочков, которые имеет сосновая пыльца, при помощи которых она и разносится ветром на большие расстояния. Срок опыления изменяется в зависимости от погоды. При ясной солнечной погоде пыльца может разлететься за 3-4 дня. В дождь этот процесс затягивается.Шишки расположены одиночно или по 2-3 штуки на опущенных вниз ножках. Незрелая шишка имеет коническую форму и темно-зеленый цвет. Иногда может присутствовать буроватый оттенок. Созревают сосновые шишки на второй год. Созревшие шишки становятся коричневого или бурого цвета. Длина составляет 3-6 см, ширина 2-3 см.
Опишите внешний вид зерна пшеницы.
Evgeni auf der flanke
Максимальная высота:
Planeta_83
У растения пшеницы – свойственный всем злакам стебель-соломина с узлами и обычно полыми междоузлиями, а листья простые, линейные, очередные, двурядные. Каждый лист отходит от узла и состоит из влагалища, охватывающего вышележащее междоузлие наподобие расщепленной трубки, и длинной узкой пластинки. На границе между влагалищем и пластинкой находятся три выроста – широкий пленчатый язычок, прилегающий к стеблю, и два охватывающих последний пальцевидных ушка. Верхнее междоузлие, или цветонос, несет соцветие – сложный колос. Он состоит из коленчатой центральной оси и очередно отходящих от нее мелких простых соцветий – колосков, обращенных к оси широкой стороной. Каждый колосок несет на своей оси от двух до пяти очередно отходящих цветков, совокупность которых прикрыта снизу двумя – верхней и нижней – колосковыми чешуями, представляющими собой кроющие листья простого соцветия. Каждый цветок защищен парой специализированных прицветников – более крупной и толстой нижней и относительно тонкой верхней цветковыми чешуями. У некоторых, т. н. остистых, сортов пшеницы нижняя цветковая чешуя оканчивается длинной остью. Цветки обычно обоеполые, с тремя тычинками и пестиком, несущим два перистых рыльца. У основания завязи находятся две или три мелкие чешуйки – цветковые пленки, или лодикулы, эквивалентные околоцветнику.
На границе между влагалищем и пластинкой находятся три выроста – широкий пленчатый язычок, прилегающий к стеблю, и два охватывающих последний пальцевидных ушка. Верхнее междоузлие, или цветонос, несет соцветие – сложный колос. Он состоит из коленчатой центральной оси и очередно отходящих от нее мелких простых соцветий – колосков, обращенных к оси широкой стороной. Каждый колосок несет на своей оси от двух до пяти очередно отходящих цветков, совокупность которых прикрыта снизу двумя – верхней и нижней – колосковыми чешуями, представляющими собой кроющие листья простого соцветия. Каждый цветок защищен парой специализированных прицветников – более крупной и толстой нижней и относительно тонкой верхней цветковыми чешуями. У некоторых, т. н. остистых, сортов пшеницы нижняя цветковая чешуя оканчивается длинной остью. Цветки обычно обоеполые, с тремя тычинками и пестиком, несущим два перистых рыльца. У основания завязи находятся две или три мелкие чешуйки – цветковые пленки, или лодикулы, эквивалентные околоцветнику.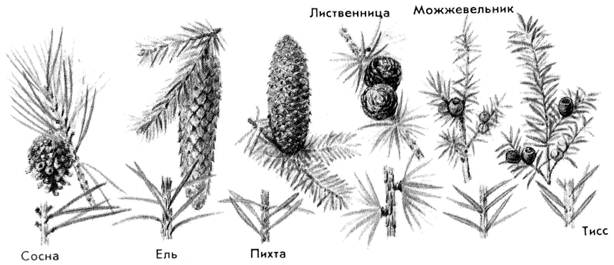 Ко времени цветения они набухают и раздвигают окружающие цветок чешуи. Пшеница – растение в основном самоопыляемое, хотя у некоторых ее типов происходит и перекрестное опыление. После оплодотворения завязь превращается в маленький твердый плод зерновку, удерживаемую в колосе цветковыми чешуями. Зерновка, или зерно, представляет собой сформировавшийся из стенки завязи околоплодник, неразрывно связанный с единственным семенем, которое содержит зародыш и эндосперм. Зародыш находится сбоку в основании зерна и состоит из почечки, корешка и прилежащей к эндосперму видоизмененной семядоли – щитка. После прорастания зародышевый корешок даст первичную корневую систему, почечка – надземные органы растения и его «взрослые» корни, а щиток будет выделять ферменты, переваривающие эндосперм, и проводить его питательные вещества к начавшему развитие всходу.
Ко времени цветения они набухают и раздвигают окружающие цветок чешуи. Пшеница – растение в основном самоопыляемое, хотя у некоторых ее типов происходит и перекрестное опыление. После оплодотворения завязь превращается в маленький твердый плод зерновку, удерживаемую в колосе цветковыми чешуями. Зерновка, или зерно, представляет собой сформировавшийся из стенки завязи околоплодник, неразрывно связанный с единственным семенем, которое содержит зародыш и эндосперм. Зародыш находится сбоку в основании зерна и состоит из почечки, корешка и прилежащей к эндосперму видоизмененной семядоли – щитка. После прорастания зародышевый корешок даст первичную корневую систему, почечка – надземные органы растения и его «взрослые» корни, а щиток будет выделять ферменты, переваривающие эндосперм, и проводить его питательные вещества к начавшему развитие всходу.
Можжевельник в отличие от сосны и ели образует не просто шишки, а так называемые шишкоягоды, которые внешне похожи на ягоду, но с точки зрения ботаники по своему строению являются шишкой.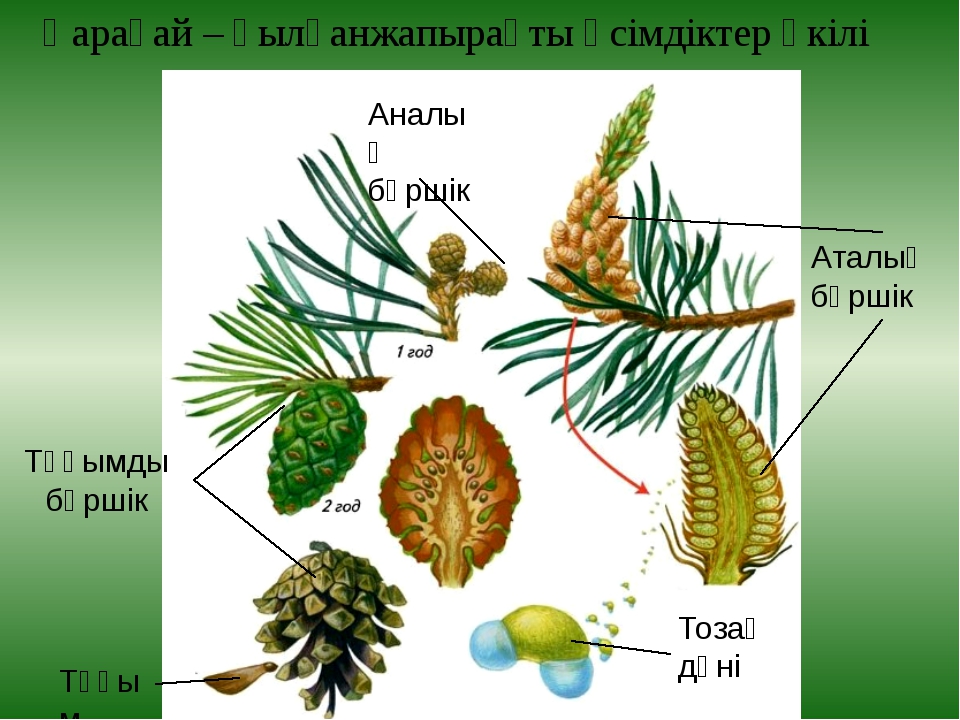 В отличие от сосны и ели женские и мужские шишки у можжевельника созревают на разных растениях. На одном растении созревают женские шишки, из которых потом образуются шишкоягоды, а на другом только мужские пыльцевые шишки. Поэтому если вы посадили у себя на участке можжевельник и хотите добиться от него плодов, то растению обязательно нужна будет пара.
В отличие от сосны и ели женские и мужские шишки у можжевельника созревают на разных растениях. На одном растении созревают женские шишки, из которых потом образуются шишкоягоды, а на другом только мужские пыльцевые шишки. Поэтому если вы посадили у себя на участке можжевельник и хотите добиться от него плодов, то растению обязательно нужна будет пара.
Нарисовать хвоинки пихты поодиночке парами пучками
Окружающий мир 4 класс 3. В атласе-определителе “От земли до неба” прочитай о хвойных деревьях (с. 32). Обрати внимание на различия их хвои. Нарисуй хвоинки, показав особенности их формы и расположения на веточках (поодиночке, парами, пучками).
В природе существует множество хвойных деревьев, есть среди них даже те, которые растут в очень жарких местах — субтропиках и тропиках. Но в нашей стране больше распространены лиственницы, ели, сосны и кедры, причем последние иногда называют кедровыми соснами.
У любого хвойного дерева есть иголки-хвоинки, которые заменяют им листья и эти хвоинки отличаются и размерами, и твердостью, и тем, как они растут на веточке.
Так у елки хвоинки короткие и жесткие, растут по одной. У пихты хвоинки такие же, только гораздо мягче и гуще.
А вот у кедра хвоинки длинные и парные, растут пучками. Парные хвинки имеет и сосна. Хвоинки лиственницы одиночные, но также растут пучками. Помня об этом давайте дорисуем хвоинки разных деревьев:
Подробное решение часть 1 (страница) Стр. 46 по окружающему миру часть 1, 2 для учащихся 4 класса, авторов А.А. Плешаков, Е.А. Крючкова 2016
- Гдз по Окружающему миру за 4 класс можно найти тут
- Гдз тесты по Окружающему миру за 4 класс можно найти тут
- Гдз контрольно-измерительные материалы по Окружающему миру за 4 класс можно найти тут
2. (с. 47) Выполните задания для работы в группах.
1) (с. 47) Узнайте хвойные деревья по веточкам и шишкам. Пронумеруйте рисунки.
1.Ель. 2.Сосна. 3.Кедровая сосна. 4.Пихта. 5.Лиственница.
Рисунки расположены в следующем порядке:
2) Узнайте лиственные деревья по листьям и плодам. Пронумеруйте рисунки.
Пронумеруйте рисунки.
1.Дуб. 2.Клен. 3.Липа. 4.Ясень. 5.Вяз.
3) (с.48) На этом рисунке «спрятались» 7 животных тайги. Найдите их и назовите. Составьте список животных, которых вам удалось отыскать на рисунке.
3. (с. 48) В атласе-определителе «От земли до неба» прочитай о хвойных деревьях. Обрати внимание на различия их хвои. Нарисуй хвоинки, показав особенности их формы и расположения на веточках (поодиночке, парами, пучками).
4. (с. 49) В книге «Зеленые страницы» прочитай о каком-либо растении или животном лесных зон (по своему выбору). Запиши 1-2 интересных факта.
Белки живут в дупле или же строят гнезда. В гнезде белки даже при минус десяти будет около +10 градусов. У белок очень хорошее обоняние.
5. (с. 49) Составь схему цепи питания, характерной для тайги. Сравни ее со схемой предложенной соседом по парте. С помощью этих схем расскажи об экологических связях в зоне тайги.
6. (с. 49) По заданию учебника (с. 102) нарисуй, как ты представляешь себе разные леса.
7. (с. 50) Здесь можешь сделать записи для своего сообщения по заданию учебника (с. 102).
Тема сообщения: Кедровка.
1)Кто такая кедровка?
2) Как выглядит птичка?
3) Где обитает кедровка?
Важная информация для сообщения:
Есть в семействе врановых птица с интересным названием – ореховка. По-другому ее еще называют кедровка. Длина тела может достигать 36 см, хвост – 11 см, на конце слегка закруглен и окаймлен светлой полоской. Масса тела варьируется от 120 до 200 г. Оперение плотное, окраска пестрая. Спинка, голова и брюшко окрашены в темно- коричневый цвет с разбросанными то тут, то там белыми пятнышками. Крылья темные, буро-коричневые, с оттенками черного цвета.
Свое имя птица получила из-за того, что питается она в основном кедровыми орехами. Поэтому и живут кедровки там, где растут кедровые леса. На таежных просторах европейской и азиатской частей России, в горных лесах Европы гнездится эта шумная, крикливая птица.
На таежных просторах европейской и азиатской частей России, в горных лесах Европы гнездится эта шумная, крикливая птица.
готовые домашние задания
1. Найдите на фотографиях лиственные и хвойные деревья. Лиственные деревья обозначьте жёлтым кружком, а хвойные — зелёным.
К лиственным деревьям относятся: тополь, липа, осина. Из обозначаем желтым кружком.
К хвойным деревьям относятся: сосна, ель и лиственница. Их обозначаем зеленым кружком.
2. Практическая работа «Определяем хвойные деревья».
1) Зарисуйте рассмотренные вами веточки, хвоинки, шишки. Рядом нарисуйте силуэт дерева, которому они принадлежат.
Вот как выглядят лиственница, сосна и ель.
А вот так выглядят хвоинки сосны, ели и лиственницы. Обратите внимание на разницу между хвоинками. У сосны хвоинки расположены по две. У ели хвоинки расположены поодиночке. У лиственницы хвоинки расположены пучками по 20-40 хвоинок в пучке.
А вот как выглядят шишки сосны, ели и лиственницы. Обратите внимание на разницу между шишками. У сосны шишка овальная. У ели шишка в форме конуса. А у лиственницы шишка округлая (можно сказать пузатая).
Обратите внимание на разницу между шишками. У сосны шишка овальная. У ели шишка в форме конуса. А у лиственницы шишка округлая (можно сказать пузатая).
А вот как вы можете нарисовать контуры ели, сосны и лиственницы.
Лиственница Ель Сосна
2) Сообщите другим ребятам о результатах своей работы. Выслушайте и оцените их сообщения.
3. Дорисуй веточки. От каких они деревьев? Объясни, по каким признакам тебе удалось это узнать
Дорисуй веточки. От каких они деревьев?
Мне удалось узнать ель и сосну по следующим признакам.
У ели длина хвоинок маленькая (около 1 сантиметра). Хвоинки растут по одной.
У сосны хвоинки длинные (около 4-5 сантиметров). Хвоинки растут парами.
4. Соберите на прогулке шишки и смастерите из них забавную фигурку. Вот несколько примеров.
Урок 41. Отдел голосеменные.
 Особенности строения и жизнедеятельности голосеменных
Особенности строения и жизнедеятельности голосеменныхМетодическое пособие разработки уроков биологии 6класс
Тип урока – комбинированный
Методы: частично-поисковый, проблемного изложения, репродуктивный, объяснительно-иллюстративный.
Цель:
– осознание учащимися значимости всех обсуждаемых вопросов, умение строить свои отношения с природой и обществом на основе уважения к жизни, ко всему живому как уникальной и бесценной части биосферы;
Задачи:
Образовательные: показать множественность факторов, действующих на организмы в природе, относительность понятия «вредные и полезные факторы», многообразие жизни на планете Земля и варианты адаптаций живых существ ко всему спектру условий среды обитания.
Развивающие: развивать коммуникативные навыки, умения самостоятельно добывать знания и стимулировать свою познавательную активность; умения анализировать информацию, выделять главное в изучаемом материале.
Воспитательные:
Формирование экологической культуры на основе признания ценности жизни во всех её проявлениях и необходимости ответственного, бережного отношения к окружающей среде.
Формирование понимания ценности здорового и безопасного образа жизни
УУД
Личностные:
воспитание российской гражданской идентичности: патриотизма, любви и уважения к Отечеству, чувства гордости за свою Родину;
Формирование ответственного отношения к учению;
3) Формирование целостного мировоззрения, соответствующего современному уровню развития науки и общественной практики.
Познавательные: умение работать с различными источниками информации, преобразовывать её из одной формы в другую, сравнивать и анализировать информацию, делать выводы, готовить сообщения и презентации.
Регулятивные: умение организовать самостоятельно выполнение заданий, оценивать правильность выполнения работы, рефлексию своей деятельности.
Коммуникативные: Формирование коммуникативной компетентности в общении и сотрудничестве со сверстниками, старшими и младшими в процессе образовательной, общественно полезной, учебно-исследовательской, творческой и других видов деятельности.
Планируемые результаты
Предметные: знать – понятия «среда обитания», «экология», «экологические факторы» их влияние на живые организмы, «связи живого и неживого»;. Уметь – определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.
Личностные: высказывать суждения, осуществлять поиск и отбор информации; анализировать связи, сопоставлять, находить ответ на проблемный вопрос
Метапредметные:.
Умение самостоятельно планировать пути достижения целей, в том числе альтернативные, осознанно выбирать наиболее эффективные способы решения учебных и познавательных задач.
Формирование навыка смыслового чтения.
Форма организации учебной деятельности – индивидуальная, групповая
Методы обучения: наглядно-иллюстративный, объяснительно-иллюстративный, частично-поисковый, самостоятельная работа с дополнительной литературой и учебником, с ЦОР.
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Цели: дать представление о семенных растениях, раскрыть преимущества семенного размножения перед споровым; показать особенности строения голосеменных растений; выявить черты сходства и различия представителей споровых и семенных растений; познакомить с особенностями размножения голосеменных растений на примере цикла размножения сосны; продолжить формирование навыков работы с натуральными объектами, выполнения биологического рисунка.
Оборудование и материалы: карточки с вопросами, живые побеги сосны и ели с шишками, шишки различных голосеменных растений, гербарные экземпляры различных голосеменных растений, таблицы: «Строение и размножение сосны», «Растения елового леса», «Растения соснового леса», коллекции шишек и побегов голосеменных растений, побеги цветковых растений, несколько моделей или натуральных плодов покрытосеменных растений, шишки сосны и ели, погрызенные белкой, поклеванные клестом или дятлом.
Ключевые слова и понятия: семенные растения, голосеменные и цветковые растения, древесные растения, кустарники, стелющиеся формы растений; эволюция, перекрестное опыление, шишка — видоизмененный побег, плод; однодомное растение, женские шишки, мужские шишки, стержень (ось), чешуи; семязачаток, яйцеклетка, эндосперм, неподвижные мужские гаметы — спермии; зигота, зародыш, семенная кожура, пыльцевые мешочки, смола; появление семян — эволюционный шаг, дающий преимущество семенным растениям.
Ход урока
Актуализация знаний
Работа у доски
(К доске вызывают 3 учеников. Каждому дают по карточке с вопросами. На выполнение задания отводится около 5 мин.)
Карточка 1
Запишите основные черты, характерные для водорослей.
Приведите примеры различных водорослей.
Карточка 2
Запишите основные черты, характерные для мохообразных.
Приведите примеры различных мохообразных.
Карточка 3
Запишите основные черты, характерные для папоротникообразных.
Приведите примеры различных папоротникообразных.
(По истечении времени учащиеся устно отвечают на поставленные вопросы. Класс сравнивает и анализирует их ответы, исправляет ошибки и неточности.)
Изучение нового материала
Рассказ учителя с элементами беседы
Сегодня мы приступаем к изучению новой группы растений — отдела голосеменные. Принципиальное отличие растений этой группы от растений, изученных нами ранее, состоит в способе размножения.
Вспомните, посредством чего размножались растения, с которыми мы уже познакомились. (Этирастения размножались посредством спор.)
Способ размножения представителей отдела голосеменные принципиально иной.
Как вы думаете, посредством чего размножаются эти растения? (Ответы учащихся.)
Они размножаются с помощью семян. Возникновение семян — очень важный этап в эволюции растений.
Отдел голосеменных насчитывает около 700 видов. Представители этой группы — в основном, деревья, реже — кустарники и стелющиеся формы растений. Трав среди голосеменных нет.
Самостоятельная работа учащихся с учебником
Пользуясь текстом учебника (учебник И.Н. Пономаревой § 42; учебник В.В. Пасечника § 16), выпишите основные признаки голосеменных растений.
На выполнение задания отводится 5—7 мин. Учащиеся по очереди называют по одному признаку растений отдела голосеменные. Учитель дополняет ответы.)
Признаки растений отдела голосеменные
Размножение посредством семян.
Не образуют плодов.
Древесные растения или кустарники, иногда стелющиеся формы.
Листья чаще всего игольчатые, слегка уплощенные или чешуевидные.
Чаще всего вечнозеленые растения.
Настоящие сосуды отсутствуют.
Разнополые растения.
Оплодотворение происходит без участия воды.
Мужские гаметы — неподвижные спермии.
Продолжение рассказа учителя с элементами беседы
Свое название голосеменные получили благодаря семенам, которые лежат открыто, как бы голо, и в отличие от семян покрытосеменных растений не защищены стенками плода.
Тем не менее наличие семян является большим эволюционным шагом, дающим этим растениям огромное преимущество перед споровыми.
Как вы считаете, в чем это проявляется? (Ответы учащихся.)
В отличие от спор в семенах имеется запас питательных веществ, необходимых зародышу в первое время его развития, пока у молодого растения не сформируется корневая система. Кроме того, внутри семени зародыш будущего растения надежно защищен от неблагоприятных воздействий окружающей среды. Все это значительно увеличивает шансы молодого растения на выживание.
Кроме того, внутри семени зародыш будущего растения надежно защищен от неблагоприятных воздействий окружающей среды. Все это значительно увеличивает шансы молодого растения на выживание.
Что представляется вам, когда речь заходит о хвойных растениях? (Ель и сосна.)
Ель и сосна — наиболее широко известные представители хвойных голосеменных растений средней полосы.
Практическая работа. СТРОЕНИЕ ХВОЙНЫХ ГОЛОСЕМЕННЫХ РАСТЕНИЙ СОСНЫ И ЕЛИ
Цели: познакомить с внешним строением хвойных голосеменных растений на примере сосны обыкновенной и ели обыкновенной, с микроскопическим строением хвои; продолжить формирование навыков работы с натуральными объектами, выполнения биологического рисунка.
Оборудование: живые или гербарные побеги сосны и ели, шишки сосны, шишки ели, готовый микропрепарат «Хвоя сосны», микроскоп.
Общие рекомендации. Перед началом работы необходимо напомнить учащимся о правилах техники безопасности.
Перед началом работы необходимо напомнить учащимся о правилах техники безопасности.
Ход работы
Рассмотрите внешнее строение побега сосны. Определите размер хвоинок, как они располагаются на побеге.
Рассмотрите внешнее строение шишки сосны. Какую форму она имеет?
Зарисуйте внешнее строение побега и шишку сосны.
Рассмотрите внешнее строение побега ели. Определите размер хвоинок, как они располагаются на побеге.
Рассмотрите внешнее строение шишку ели. Какую форму она имеет?
Зарисуйте внешнее строение побега и шишку ели.
Рассмотрите готовый микропрепарат «Хвоя сосны» под микроскопом при малом и большом увеличении.
Найдите кожицу, выполняющую защитные функции, устьица. Как расположены устьица? Подсчитайте примерное количество устьиц.
Зарисуйте микроскопическое строение хвои сосны.
Сделайте вывод о чертах сходства и чертах различия сосны и ели, а также о приспособленности этих растений к жизни в суровом климате.
Сравнительную характеристику сосны и ели
Признак | Сосна обыкновенная | Ель обыкновенная |
Характер хвои | Хвоинки длинные, 3—5 см в длину, расположены попарно на сильно укороченных побегах | Хвоинки короткие (1,5—2 см), расположены одиночно, густо покрывают побеги |
Расположение веток | Ветви собраны в мутовки, по которым можно примерно определить возраст дерева. | Ветви распределены от верхушки до основания |
Шишки | Относительно округлые | Более вытянутые, с гладкими чешуями |
Характер корневой системы | Пластичная корневая система. Зависит от условий произрастания дерева | Главный корень обычно развит плохо |
Требования к освещению | Светолюбивая порода. Образует светлые леса (боры) | Тенелюбивая порода. Образует темнохвойные леса |
 Нижняя часть ствола не имеет веток
Нижняя часть ствола не имеет ветокТребования к почве | Неприхотлива по отношению к почвам | Требовательна к субстрату и минеральному питанию |
Потребность во влаге | Нетребовательна по отношению к влаге | Влаголюбива |
Опыление и распространение семян | Ветроопыляемое растение. | Ветроопыляемое растение. Семена распространяются животными и ветром |
 Семена распространяются животными и ветром
Семена распространяются животными и ветромПродолжение рассказа учителя с элементами беседы
Как мы уже говорили, голосеменные растения (например, сосна и ель) размножаются семенами.
Где образуются семена у этих растений? (В шишках.)
А как размножаются цветковые растения? (Семенами.)
В чем же состоит основное отличие семенного размножения покрытосеменных и голосеменных растений? (Ответы учащихся.)
Покрытосеменные растения образуют плод, внутри которого образуются и созревают семена, а у голосеменных растений семена не прикрыты стенками плода. Они расположены в шишках, на чешуях, т. е. семена как бы голые, отсюда и название отдела.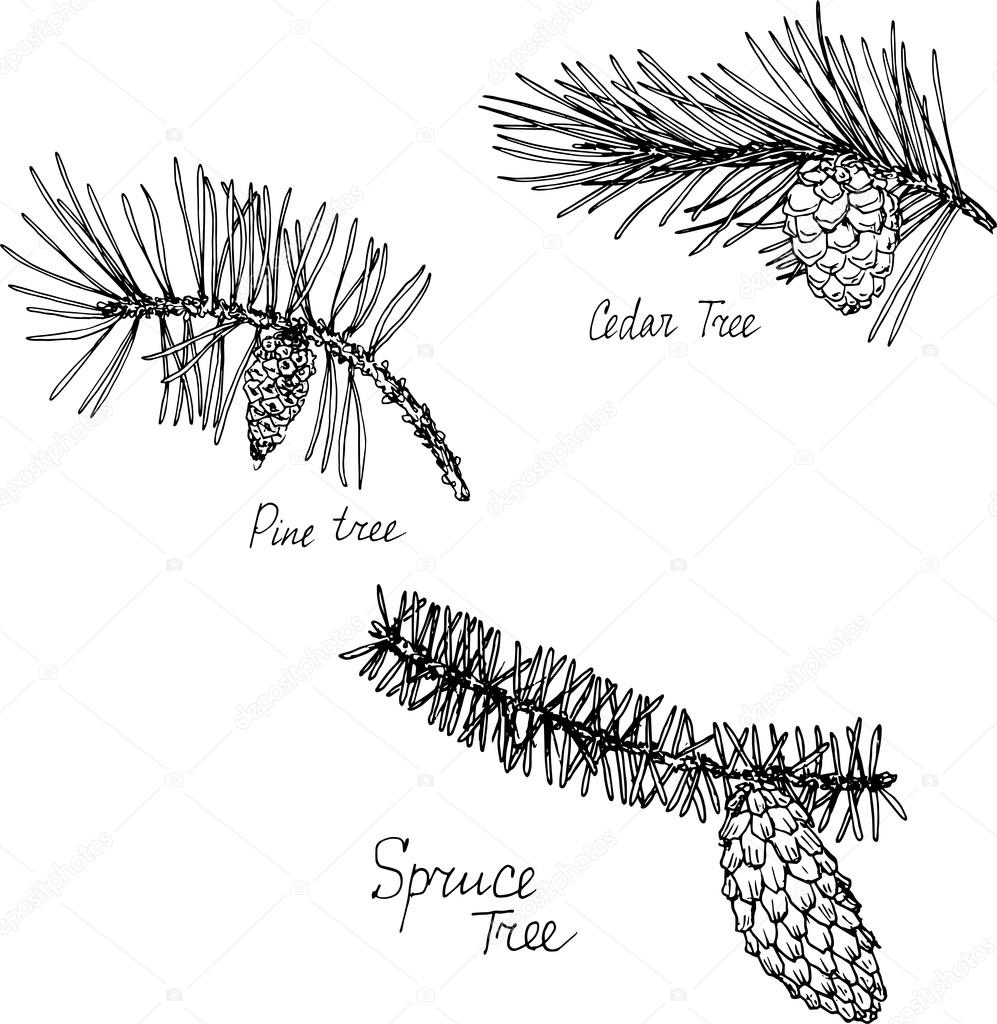
Давайте более подробно рассмотрим цикл развития сосны. (Учитель сопровождает свой рассказ демонстрацией гербариев, побегов растений, таблиц.) Сосна, так же как и ель, однодомное растение.
Вспомните, что значит «однодомное». (Это значит, что мужские и женские гаметы развиваются на одном растении.)
Плодоносить эти растения начинают на 15-30-м году жизни. На верхушках молодых побегов сосны в конце весны — начале лета можно увидеть небольшие красноватые шишечки. Это молодые женские шишки. Шишка представляет собой видоизмененный побег. У шишки можно выделить стержень (или ось), на котором расположены чешуи — видоизмененные листья. На верхней стороне каждой из чешуй расположено по два семязачатка.
Мужские шишки более мелкие, всего 1—2 см в длину, желтоватые, собраны в небольшие тесные группы. Мужские шишки растут ниже женских на тех же или на других ветках.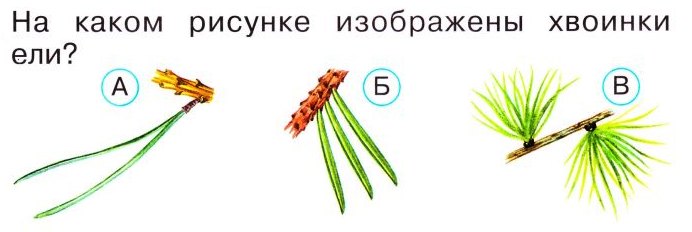 На нижней стороне чешуек шишки размещено по 2 пыльцевых мешочка, в которых созревает пыльца. Каждая пыльца сосны имеет по два небольших пузырька, наполненных воздухом.
На нижней стороне чешуек шишки размещено по 2 пыльцевых мешочка, в которых созревает пыльца. Каждая пыльца сосны имеет по два небольших пузырька, наполненных воздухом.
Как вы думаете, для чего это нужно? (Ответы учащихся.)
Благодаря наличию этих пузырьков пыльца очень легкая
и легко переносится ветром.
Каков биологический смысл расположения мужских шишек ниже, чем женских? (Ответы учащихся.)
Это приспособление к перекрестному опылению. Пыльца, высыпаясь из шишек, с большей вероятностью попадет не на шишки, расположенные на том же дереве или той же ветке, а на шишки другого дерева.
После того как пыльца сосны попадает в семязачатки (происходит опыление), чешуйки смыкаются и склеиваются смолой. Но оплодотворения пока что не происходит. Семязачатки в это время еще не готовы к оплодотворению.
Через год женские шишки становятся зелеными, увеличиваются в размерах. Внутри семязачатков созревают яйцеклетки и образуется питательная ткань — эндосперм. После созревания семязачатков происходит оплодотворение неподвижными мужскими гаметами — спермиями.
Внутри семязачатков созревают яйцеклетки и образуется питательная ткань — эндосперм. После созревания семязачатков происходит оплодотворение неподвижными мужскими гаметами — спермиями.
С какой группой растений это роднит голосеменные? (Подобные мужские гаметы характерны для покрытосеменных растений.)
После оплодотворения образуется зигота, из которой прямо на материнском растении начинает развиваться зародыш, который питается за счет эндосперма. В результате образуется семя, состоящее из зародыша, питательной ткани эндосперма и семенной кожуры. Через 1,5 года шишка сосны высыхает, трескается и семена высыпаются на поверхность земли.
Семена голосеменных растений распространяются животными или ветром. Они имеют особое пленчатое крылышко, способствующее перемещению их при помощи ветра. (Учитель демонстрирует семена сосны.) Семена сосны и ели часто поедают птицы и белки. Часть семян при этом высыпается и прорастает. (Учитель может продемонстрировать шишки сосны и ели, объеденные белкой или поклеванные клестом или дятлом.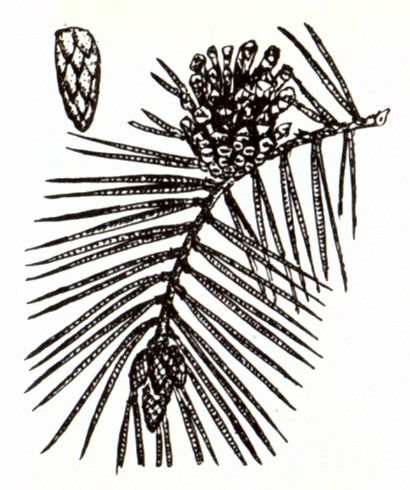 ) Семя намного тяжелее одноклеточной споры, но гораздо лучше защищено от неблагоприятных воздействий окружающей среды, кроме того, снабжено запасом питательных веществ. При попадании в благоприятные условия семена прорастают и дают начало новому организму.
) Семя намного тяжелее одноклеточной споры, но гораздо лучше защищено от неблагоприятных воздействий окружающей среды, кроме того, снабжено запасом питательных веществ. При попадании в благоприятные условия семена прорастают и дают начало новому организму.
Подсчитайте, сколько времени необходимо такому голосеменному растению, как сосна, для прохождения полного цикла развития. (Ответы учащихся.)
Закрепление знаний и умений
Фронтальный опрос
Ответьте на вопросы.
Какие растения называют голосеменными?
В чем состоит принципиальное отличие голосеменных растений от споровых?
Каковы основные признаки голосеменных растений?
Где развиваются семена голосеменных растений?
В чем заключается отличие семян голосеменных растений от семян покрытосеменных?
Каково строение женской и мужской шишек сосны?
В чем отличие в строении этих шишек?
Какие особенности строения возникли у пылинок сосны, которые способствуют их переносу ветром?
Как происходит опыление и оплодотворение у голосеменных растений (на примере сосны)?
Какими способами распространяются семена сосны?
В чем состоит эволюционное преимущество семенных растений перед споровыми?
Самостоятельная работа учащихся в парах
(Необходимо составить 2—3 тестовых вопроса по теме урока с 4 вариантами ответов, один из которых является правильным. Вопросы аккуратно пишут на отдельном листе бумаги. На выполнение задания отводится 5—7 мин. После чего учащиеся отвечают на вопросы соседа, оценивая при этом корректность и правильность выполненного задания, и наоборот.)
Вопросы аккуратно пишут на отдельном листе бумаги. На выполнение задания отводится 5—7 мин. После чего учащиеся отвечают на вопросы соседа, оценивая при этом корректность и правильность выполненного задания, и наоборот.)
Подведение итогов урока
Домашнее задание
Прочитать параграф, знать основные термины, устно ответить на вопросы в конце параграфа. (Учебник И.Н. Пономаревой § 42; учебник В.В. Пасечника § 16.)
Творческое задание. Узнать, какие животные питаются семенами голосеменных растений. Попробовать найти в лесу шишки, которые ели белки, клесты, дятлы или другие животные. Чем внешне отличаются эти шишки? Изобразить шишки и животных, которые ими питались.
Задания для учеников, интересующихся биологией.
Нарисовать графическую схему, отражающую цикл развития сосны.
Рассмотреть живые побеги и изображения различных голосеменных растений вашей местности. Выписать признаки, характерные только для конкретного растения, т. е. те, по которым это растение можно отличить от других голосеменных. Зарисовать отдельные побеги и шишки этих растений. Можно составить мини-определитель хвойных растений, встречающихся в вашей местности.
Выписать признаки, характерные только для конкретного растения, т. е. те, по которым это растение можно отличить от других голосеменных. Зарисовать отдельные побеги и шишки этих растений. Можно составить мини-определитель хвойных растений, встречающихся в вашей местности.
Подготовить доклад об интересных представителях голосеменных растений.
Голосеменные
Размножение голосемянных
Отдел Голосеменные. Общая характеристика.AVI
 youtube.com/embed/icTEoYNL3dg”>
youtube.com/embed/icTEoYNL3dg”>
Голосеменные-лекция
Ресурсы:
И.Н. Пономарёва, О.А. Корнилова, В.С. Кучменко Биология : 6 класс : учебник для учащихся общеобразовательных учреждений
Серебрякова Т.И., Еленевский А. Г., Гуленкова М. А. и др. Биология. Растения, Бактерии, Грибы, Лишайники. Пробный учебник 6—7 классов средней школы
Н.В. Преображенская Рабочая тетрадь по биологии к учебнику В В. Пасечника «Биология 6 класс. Бактерии, грибы, растения»
В.В. Пасечника. Пособие для учителей общеобразовательных учреждений Уроки биологии. 5—6 классы
Калинина А.А. Поурочные разработки по биологии 6класс
Вахрушев А. А., Родыгина О.А., Ловягин С.Н. Проверочные и контрольные работы к
А., Родыгина О.А., Ловягин С.Н. Проверочные и контрольные работы к
учебник «Биология», 6-й класс
Биоуроки http://biouroki.ru/material/lab/2.html
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
– http://ppt4web.ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
Защита хвойных деревьев от стволовых вредителей
Весной складываются благоприятные условия для массового распространения стволовых вредителей хвойных деревьев, в связи с чем наблюдается частое поражение молодых посадок елей, сосен и туи. На пораженных растениях можно наблюдать небольшие отверстия – ходы на стволе, из повреждений начинает выделяться смола, в результате чего ствол покрывается смоляными потеками, меняется цвет хвои с ярко-зеленого на бледный, летом можно наблюдать появление опилок на стволе и под деревом, также можно услышать потрескивание – результат работы жука.
Стволовые вредители – большая группа насекомых, питающихся тканями ствола дерева. Под корой и в древесине ствола и ветвей они прогрызают ходы, имеющие характерную для каждого вида форму. Это личинки жуков семейства короедов, усачей, златок, а также личинки перепончатокрылых из семейства рогохвостов. Меньшее значение имеют сверлильщики, точильщики и др.
Личинки короедаКороедУсачПоражению подвержены прежде всего ослабленные деревья, а также молодые посадки, пока растения приживаются и адаптируются на новом месте. Особое внимание молодым посадкам надо уделять в местах, где рядом располагаются леса с хвойными породами, откуда может происходить массовый лет жуков-вредителей. Для посадки в таких местах лучше выбирать не копаные, а здоровые контейнерные растения с хорошей корневой системой, которые можно пересадить без повреждения корней и существенного ослабления растений.
Массовый лет жуков наблюдается в мае-июне. Цикл развития личинок проходит в течение 1,5-2 месяцев. У некоторых видов наблюдается повторный лет в июле-августе. В это время необходимо проводить профилактические обработки для защиты деревьев. При появлении первых признаков заражения необходимо срочно принимать меры, иначе пораженные растения быстро усыхают и при массовом поражении погибают в течение одного сезона.
Цикл развития личинок проходит в течение 1,5-2 месяцев. У некоторых видов наблюдается повторный лет в июле-августе. В это время необходимо проводить профилактические обработки для защиты деревьев. При появлении первых признаков заражения необходимо срочно принимать меры, иначе пораженные растения быстро усыхают и при массовом поражении погибают в течение одного сезона.
Для защиты растений от стволовых вредителей применяют системный препарат АКТАРА. В профилактических целях проводят опрыскивание в период массового лета жуков (май-июнь, конец июля-август раз в две недели) раствором препарата 10 г. на 10 литров воды из опрыскивателя по кроне растения и по стволу до полного смачивания всей поверхности. При первых признаках поражения растений дополнительно проводят полив под корень раствором препарата из расчета 30 г. на 10 литров воды, ведро раствора на 1 растение. Также можно делать инъекции препарата высокой концентрации из шприца непосредственно в отверстия на стволе дерева. Также необходимо обеспечить требуемый уход за пересаженными растениями для их быстрой приживаемости и повышения естественного уровня сопротивляемости вредителям.
Также необходимо обеспечить требуемый уход за пересаженными растениями для их быстрой приживаемости и повышения естественного уровня сопротивляемости вредителям.
Правила обработки хвойных растений
- Для уничтожения насекомых и личинок используйте современные инсектициды, при обработке от клеща – акарициды, против грибковых заболеваний – фунгициды. Все препараты можно найти в нашем магазине товаров для сада либо в других аналогичных.
- При обработке всегда соблюдайте прилагаемые инструкции препаратов и самостоятельно не экспериментируйте со смешиванием разных препаратов и увеличением дозировки.
- Обрабатывайте хвойные растения такие как ель, сосна и туя ранним утром или вечером в сухую погоду. Приготовленный раствор должен впитаться в опрыскиваемый куст или дерево, поэтому в жару или пасмурную погоду (дождь) не стоит проводить обработку.
- Старайтесь проводить обработку систематично если заметили появление молодых и новых вредителей (личинок), скорее всего ранее паразиты уже успели отложить яйца, а грибки – споры. Чередуйте препараты, так как одному и тому же препарату может появиться резистентность (привыкание).
- Обязательно используйте средства индивидуальной защиты при обработке хвойников, не проводите обработку в ветреную погоду и изолируйте на время домашних питомцев.
 Чередуйте препараты, так как одному и тому же препарату может появиться резистентность (привыкание).
Чередуйте препараты, так как одному и тому же препарату может появиться резистентность (привыкание).Эти не хитрые советы помогут вам в будущем избежать лишней потери времени, средств и гибели любимых растений.
Вредители хвойных и меры борьбы
| Симптомы поражения | Причина | Меры борьбы |
|---|---|---|
| Мелки насекомые жёлто-зелёного цвета опушённые, сидят на побегах между хвоинками. Блестящие насекомые на ветке и коре | Опушённая либо корьевая тля | Используем для обработки инсектициды |
| На коре или хвое появились белоснежные пушистые и липкие образования под которыми мелкие черные насекомые | Хермес или корьевой хермес | Также используем системные инсектициды |
| Мелкие гусеницы зелёного цвета, поедают хвою и обвивают её паутиной | Пяденицы, листоверкти, моль | Применяем инсектициды согласно инструкции |
| Небольшие образования в виде лепёшки белого или темно-коричневого цвета под которой находятся личинки | Ложнощитовка | Инсектициды либо применяем карбофос, актеллика и подобные препараты |
| Между ветвями множество тонких паутинок, внутри которых передвигаются мелкие насекомые, хвоя буреет | Паутинный клещ | Используем акарициды |
Заболевания хвойных и меры борьбы
| Симптомы поражения | Причина | Меры борьбы |
|---|---|---|
| На ветвях и хвое веретенообразные вздутия с слизистыми выделениями | Ржавчина |
Обрезаем пораженные ветви либо уничтожаем полностью дерево если поражена большая его часть. Дезинфицируем и обрабатываем раны фунгицидами Дезинфицируем и обрабатываем раны фунгицидами
|
| Хвоя после таяния снега буреет и опадает. Страдают молодые хвойные растения обычно | Шютте обыкновенное | Обрабатываем фунгицидами, применяется обычно бордоская смесь или коллоидная сера, удаляем пораженную хвою |
| Вертикальное растрескивание коры, часто встречается на молодых туях и кипарисовиках | Обморожение коры | Утепляем деревья на зиму, раны мажем садовым варом, обматываем или стягиваем кору до срастания |
| Побурение и осыпание хвои с одной стороны кустарника или дерева | Весенний солнечный ожог | Затеняем растение с солнечной стороны ранней весной |
| Раковые язвы на ветвях и стволе, на отпавшей коре образуются грибковые белые выделения | Ступенчатый рак | Удаляем погибшие ветви или растения |
В любом случае, если чувствуете, что самостоятельно не можете определить причину и методы борьбы с вредителями и болезнями хвойных, то лучше обратиться к специалистам и агрономам, так вы получите точные рекомендации или гарантии.
Ознакомьтесь с полезной статьей зачем укрывать хвойные растения на зиму и когда снимать укрытие не навредив, а также чем укрывать и как.
К спискуЧем отличается ель от сосны, в чем разница? Разница между елью и сосной
Какое дерево выбрать, если вы хотите вырастить собственную «елку»? Разговаривая о деревьях, мы нередко упоминаем некую «елку» как собирательный образ для хвойного растения конической формы с пахучей хвоей. Но как понять, о каком растении на самом деле идет речь: о ели или о пихте? Давайте учиться их различать!
Ель и пихта: что общего?
Говоря о ели и пихте, обычно подразумевают самые распространенные виды – ель обыкновенную и пихту сибирскую . Именно их чаще всего можно встретить в лесах, лесопарках и городском озеленении. Интересно смотрятся , в том числе из разных сортов ели и пихты.
Ель и пихта имеют достаточно много общих признаков:
- конусовидная крона,
- высота взрослого дерева (около 30 м),
- раскидистые лапы-ветви почти от самой земли,
- хвоя,
- продолжительность жизни.
.jpg)
Эти виды достаточно просто отличать, зная некоторые секреты. Шишки, хвоя и ствол у этих деревьев совершенно разные.
____________________________________________________________
Фото: ель обыкновенная
Ель и пихта: как отличить?
Таблица: отличия ели и пихты
Ель и пихта: что выбрать?
Если речь идет о выборе между елью обыкновенной и пихтой сибирской , как наиболее ярких представителей своего вида, то для посадки в саду, пожалуй, стоит отдать предпочтение пихте. Кора у нее более привлекательная, крона густая, насыщенная и очень декоративная.
В качестве новогоднего дерева лучше выбрать пихту, т. к. она даст больше аромата
_____________________________________________________________
Фото: пихта сибирская
У родов пихты и ели существует огромное количество сортов с разнообразной окраской хвои, формой кроны и высотой.
При выборе можно полагаться на:
- условия произрастания,
- основы композиции,
- личные предпочтения.
_______________________________________________________________
Знания отличий древесины сосны от ели дают возможность сделать правильный выбор относительно материала для строительства бани, деревянного дома или производства пиломатериалов. Итак, далее будут рассмотрены особенности древесины сосны и ели:
Отличия сосны от ели:
1) Две данные породы отличаются однородностью, так как в сосне имеются более ярко выраженные смоляные ходы. Такая однородность обеспечивает ели небольшое преимущество в теплоизоляционных свойствах.
2) Древесина сосны и ели различаются по внешнему виду, а именно по выраженному рисунку и цвету. Сосна обладает ярко выраженным узором и розоватым оттенком, который с годами становится еще насыщенней. У ели ситуация обратная – рисунок слабо выражен, а цвет древесины светлый с легким желтоватым оттенком. Для внутренних работ многие предпочитают использовать древесину ели из-за ее нейтральности. Однако поклонников использования сосны для внутренней отделки тоже немало.
Для внутренних работ многие предпочитают использовать древесину ели из-за ее нейтральности. Однако поклонников использования сосны для внутренней отделки тоже немало.
3) Прочность двух пород также отличается. Так как ель более рыхлая, она имеет меньший запас прочности, чем древесина сосны. Данное отличие весомо склоняет строителей на сторону сосны при возведении серьезных конструкций.
4) Вес древесины также является важным показателем, который нельзя упускать из виду. Древесина ели более легкая, чем древесина сосны.
5) В связи с особенностью структуры двух пород, следует уделить особое внимание коэффициенту поглощения влаги, который у ели значительно выше, что делает ее менее конкурентноспособной породой для наружных работ по сравнению с сосной .
6) Несмотря на более высокий коэффициент поглощения влаги у ели, она дольше сосны пропитывается антисептиком, что влечет к более продолжительной затрате рабочей силы.
7) Произрастание данных пород отличается небольшими особенностями – ель более прихотливая к почве, корневая система более поверхностна, чего нельзя сказать о сосне, которая произрастает в любом типе почвы, проникая вглубь развитой корневой системой.
8) Для ценителей аромата древесины также будет нелишним узнать об особенностях этих пород, так как в сосне более развиты смоляные, ее древесина даже покрытая лаком, благоухает годами. Ель же имеет более легкий непродолжительный и утонченный аромат.
Сходства сосны и ели:
1) Ель и сосна относится к одной и той же породе древесины – хвойные.
2) Основные характеристики двух данных пород, которые выражаются в процентном содержании целлюлозы и лигнина, говорят о большой схожести этих пород.
3) В связи со схожими характеристиками древесины ели и сосны, цена на них практически всегда одинаковая, за исключением тех случаев, когда недобросовестные торговые точки стараются нажиться на популярности сосны .
На сегодняшний день сосна является самой популярной породой для строительства большинства конструкций. Прежде всего, это связано с рядом основных характеристик, которые имеют положительные показатели. В свою очередь сосна не является дорогостоящей и капризной породой, в связи с этим цена подогревает всеобщий интерес. Если сравнивать вышеперечисленные характеристики сосны и ели, то можно сделать вывод о том, что ель замечательно подходит для внутренней отделки, так как имеет нейтральный внешний вид, тонкий ненавязчивый аромат и легкий вес.
Если сравнивать вышеперечисленные характеристики сосны и ели, то можно сделать вывод о том, что ель замечательно подходит для внутренней отделки, так как имеет нейтральный внешний вид, тонкий ненавязчивый аромат и легкий вес.
Сосна в свою очередь больше подходит в качестве основного материала конструкции, либо внешней отделки. Поэтому покупайте в качестве основного материала сосну, а в качестве отделочного – ель. Незамедлительную покупку древесины ели и сосны, можно сделать на сайте нашей торговой компании.
Большинство из нас знают такое Новогоднее дерево, как Ёлка, не задумываясь над тем, каково же настоящее имя этой красавицы. Чаще всего это ель или сосна, а точнее, ботанические названия рода, к которому каждая из них принадлежит. В наших географических условиях встречается два вида сосна обыкновенная и ель обыкновенная (иначе, европейская). Именно о них и пойдет речь.
Ель обыкновенная – одно из самых крупных хвойных деревьев, средняя высота которых 25 – 30 м . Нередко дорастает до 50 м . Форма кроны конусовидная на протяжении всего жизненного цикла. Строение корневой системы повторяет особенность наземной части дерева. Основная масса корней расположена поверхностно, именно поэтому ель склонна к ветровальности.
Нередко дорастает до 50 м . Форма кроны конусовидная на протяжении всего жизненного цикла. Строение корневой системы повторяет особенность наземной части дерева. Основная масса корней расположена поверхностно, именно поэтому ель склонна к ветровальности.
Кора взрослого растения гладкая, серого цвета. Хвойный покров образуют одиночные «иголки», которые вырастают из пазушных чешуек. Молодые шишки появляются в мае, и сначала направлены вверх. При созревании они поворачиваются и принимают висячее положение. Ель обыкновенная относится к медленнорастущим деревьям. Первые семена появляются в двадцати летнем возрасте. Урожаи повторяются в среднем один раз в 5 лет.
Сосна обыкновенная считается деревом со средней быстротой роста. В возрасте деловой древесины диаметр ствола может достигать 1 м . Высота взрослой сосны – 25 – 40 м . Срок биологической жизни около 200 лет .
Ветки молодого дерева образуют правильный геометрический конус, со временем крона постепенно приобретает вид ассиметричного зонтика. Чешуйчатая кора с глубокими трещинами имеет толщину больше 1 см. Так природа предусмотрела защиту дерева от перепада температуры и палящего солнца. Сдвоенные хвоинки, длиной 5 – 9 мм, густо расположены на одномутовчатых побегах. Созревание семян происходит на второй год после опыления.
Сходные черты
Если хорошо постараться, то можно найти сходные характеристики сосны обыкновенной и ели европейской:
- Оба представителя относятся к семейству сосновых, классу хвойные.
- Имеют видоизмененные листья в виде хвои.
- Особенность шишек в том, что они сначала растут строго вертикально, затем повисают.
- Принадлежат к деревьям первой величины свыше 20 м в высоту.
- Относятся к ветровальным деревьям.
- Выделяют бактерицидные вещества – фитонциды.
- Применяются в промышленных целях для строительства.
- В медицине используются как лекарственные средства в виде масел, настоев и отваров.
- Служат декоративным украшением в Новогодние праздники.
Не смотря на некоторое сходство отличий между ними все же больше.
Категорические противоположности
С ботанической точки зрения эти виды, совершенно разные по внешнему облику и особенностям роста:
- Форма кроны у них сходна только в юном возрасте . И если у Ели она сберегается на протяжении всей жизни, то сосна с возрастом меняет форму до неузнаваемости. Из правильного геометрического конуса образуется раскидистый беспорядочный навес.
- Быстрота роста сосны выше, чем у ели . Первая в молодом возрасте дает прирост, в среднем 50 см. ель до 10 лет растет крайне медленно.
- Структура и окраска коры различается радикально. У Ели – она тонкая, гладкая, серая. Сосновый «покров» отличается толщиной, слоистостью, цветом. Окраска в нижней части ствола — серовато бурая и намного темнее, чем на верхних ветках. Сосновая кора один из самых популярных декоративных материалов в ландшафтном дизайне для фигурного мощения.
- Еловая хвоя наполняет побеги одиночными «иголками», в то время как, на сосне образуются сдвоенные хвоинки. Их длина в два раза больше еловых.
- Семена у сосны начинают появляться на девятый год. Репродуктивная способность Ели наступает в возрасте 20 лет .
- Еловые шишки, размером 10 – 15 см, созревают на протяжении текущего года. Сосновые плоды поспевают через 18 месяцев после опыления.
- Оба вида требуют высокой аэрации грунта, не переносят уплотнения и вытаптывания.
- Обладают высокой морозоустойчивостью.
С точки зрения экологических условий, выбор места произрастания отличается радикально:
- Сосна – типично светолюбивое растение , которая чувствует себя тем лучше, чем больше солнца. Ель, напротив, предпочитает полутень и страдает в палящий зной.
- Обе культуры влаголюбивы, но в отличие от сосны, ель совершенно не переносит засухи.
- И та другая ветровальные деревья, но при сильных порывах на Сосне в первую очередь ломается крона и верхняя часть ствола. ель сразу выворачивает с корнем.
- Для полноценного роста Ели необходима богатая, плодородная рыхлая почва. Сосна обходится сравнительно бедным песчаным грунтом. Хотя, оба вида интенсивно подкисляют почву вокруг себя продуктами своей жизнедеятельности.
- Зимостойкость Ели намного меньше, чем Сосны. Для первой особенно опасны перепады ночной и дневной температуры в конце зимы, начале весны, тогда Ели нужно защищать светлым накрытием, например, агроволокном.
- Устойчивость к городской загазованности у Ели выше, чем у сосны.
- Сосну, в отличие от Ели высаживают в ветрозащитных насаждениях из-за быстроты роста.
Светлана Востропятова
Конспект НОД по познавательному развитию «Сравнение сосны и ели»
Муниципальное дошкольное образовательное учреждение «Детский сад №5»
Конспект Нод
по познавательному развитию
«Сравнение ели и сосны »
Подготовила и провела :
Востропятова С. Е.
Саратов 2018
«Сравнение ели и сосны » в старшей группе.
Форма проведения НОД : экскурсия в хвойный лес.
Цель :
Систематизировать знания детей о хвойных деревьях : ели и сосны .
Задачи :
Продолжать знакомить детей с зимним лесом
Уточнить знания о том, что сосна и ель – хвойные , вечнозеленые деревья.
Развивать умение сравнивать , находить различия и сходства между деревьями в строении стволов, расположения веток и различии хвои, шишек и семян.
Развивать интерес к исследовательской деятельности, умение находить и открывать для себя неизвестное в знакомых объектах, развивать органы чувств , обоняния, осязания и зрительное восприятие
Воспитывать бережное отношение к объектам природы и желание сохранять их живыми в ней.
Продолжать учить детей отвечать разными типами предложений
Активизация словаря : вечнозеленое, хвойные, ельник, сосновый бор , смола, колючесть, клейкость.
Предварительная работа : наблюдали деревья на участке, учувствовали в акции «Новый год без топора» , рассматривали шишки деревьев.
Подготовка воспитателя : продумала методику и содержание конспекта , подобрала иллюстративный материал, подготовила ветки деревьев как раздаточный материал.
Организация и методика проведения НОД :
Воспитатель :
Собрались все дети в круг.
Я твой друг и ты мой друг!
Крепко за руки возьмемся
И друг другу улыбнемся.
Воспитатель : Ребята я предлагаю вам отправиться со мной на экскурсию в зимний лес. Но для этого нам с вами надо сказать волшебные слова. Вы готовы?
Дети : Да
Сейчас сядем, отдохнём, и потом опять пойдём.
(дети садятся на стульчики)
(На экран воспроизводится заставка «Зимний лес» )
Воспитатель : Посмотрите ребята, как красиво в этом зимнем лесу.
Пусть сосны и ели
Всю зиму торчат
В снега и метели
Закутавшись спят
Воспитатель : Ребята, а вы узнали какие деревья стоят в снежном уборе?
Дети : Сосна и ель .
Воспитатель : Правильно, по каким признакам вы узнали эти деревья?
Дети : У сосны ствол толстый , гладкий, внизу нет веток. У елки ветки растут до самого низа. Цвет коры у сосны светлее , у ели намного темнее.
Воспитатель : Подумайте, почему у сосен ветки растут всегда в верхней части ствола, а у елей – до самого низа?
Дети : Сосна любит солнышко , а елочка нет.
Воспитатель : Верно. Сосна любит расти на солнце, поэтому все ветки она подставляет солнышку. Если сосна растет в лесу , она всеми ветками и хвоинками тянется к солнцу. Нижние ветки, оставшись в тени, без солнца, погибают, елочка же наоборот, любит расти в тени, ветки у нее расположены по всему дереву, чем ниже – тем они длиннее.
Ребята скажите, а на каком празднике мы встречаем эти деревья?
Дети : Новый год.
Воспитатель : Правильно, эти деревья украшают наши дома в этот замечательный праздник. Но как мы можем сохранить эти деревья и не остаться на Новый год без главного украшения – елочки?
Дети : Наряжать искусственную елку.
Воспитатель : Правильно, молодцы.
Воспитатель : Ребята, ель очень красивое дерево, а сосна просто уникальна . Сосна – это целая аптека , используемая человеком с древних времен. Сосна выделяет особые вещества – фитоциды, которые губительны для микробов. И что бы в этом убедиться, можно понюхать веточку сосны . Запах очень полезен, поэтому полезно, особенно в теплую погоду после дождя и грозы погулять в сосновом бору . Воздух чистый, здоровый и полезный.
Так же из сосны получают большое количество различных продуктов : сосновое масло , скипидар, канифоль, деготь и древесный уголь. Молодые побеги сосны заготавливают , сушат, а отвары и настои используют как витаминное средство при заболевании органов дыхания, полости рта и т. п.
Сосна полезна и нужна не только человеку. Как вы думаете, с кем сосна еще дружит ?
Дети : Белка, птицы, лиса, мышь, дятел и т. д.
Воспитатель : Правильно, а так же и у ели есть друзья.
Воспитатель : Ребята посмотрите, что я нашла! Корзину, давайте посмотрим, что в ней лежит? Ребята я вам хочу показать гнездо клеста, это единственная птица, которая выводит своих птенцов зимой, так как только зимой есть корм для этих птиц (показ гнезда) . Ребята скажите, а можно забирать гнезда птиц домой?
Дети : Нет
Воспитатель : Правильно, нельзя забирать гнезда из лесу. Поэтому мы гнездо оставим в лесу, а заберем с собой корзину с ветками. Теперь нам пора с вами возвращаться. (дети встают со стульев)
Мы шагаем по сугробам, по сугробам крутолобым.
Поднимай повыше ногу, проложи другим дорогу.
Очень долго мы шагали, наши ноженьки устали.
Воспитатель : Ну, вот мы и вернулись с вами обратно с группу, а теперь я предлагаю вам сесть за стол и сравнить ветки ели и сосны .
Раздаю веки
Воспитатель : У вас на столах ветки сосны и ели . Покажите мне сосновую ветку (дети поднимают ветку сосны ) . А теперь, покажите еловую (дети показывают) . Рассмотрите обе ветки внимательнее и скажите, как вы узнали их.
Ребенок : Сосну я узнала по длинным иголкам.
Ребенок : И веточка у сосны толще , чем у ели.
Ребенок : У сосны иголки растут пучками, а у елки вокруг веточки.
Ребенок : Иголки у сосны растут по 2 из одного места, а у елки – по одной.
Ребенок : У сосны иголки длиннее , а у ели – короче.
Ребенок : Ветка елочки колючая.
Ребенок : Почки на конце сосновой ветки растут , у елки тоже есть.
Ребенок : У сосны почки большие , их сразу видно, а у елки почки маленькие.
Воспитатель : (показывает ветки сосны и ели с шишками ) . У детей на столах лежат сосновые и еловые шишки . Сравните шишки сосны и ели .
Ребенок : Еловая большая, сосновая – маленькая .
Ребенок : Еловая шишка колючая, сосновая – нет .
Ребенок : Шишка еловая овальной формы, а сосновая – круглой .
Воспитатель : В шишках находятся семена и если по ним немного постучать, то семена, которые созрели, сами вылетят из шишки.
Вы хорошо рассмотрели и правильно рассказали о ветках ели и сосны . Сказали, чем ветки похожи и в чем их различие. Сосна и ель не похожи на другие деревья, такие как береза, липа и др. Как вы думаете, чем сосна и ель отличаются от них?
Дети : У сосны и ели нет листьев .
Воспитатель : Кто с этим не согласен?
Ребенок : Иголки – это их листики.
Воспитатель : Правильно, иголки – это листья. Кто знает, как правильно называются иголки сосны и ели ?
Ребенок : Хвоя.
Воспитатель : Верно, листья сосны и ели называют хвоей. Поэтому эти деревья называют хвойными. Положите веточку себе на ладошку, накройте ладонью и слегка потрите. Что почувствовали?
Дети : Пахнет елкой.
Воспитатель : Из хвойных деревьев добывают эфирные масла (показать, которые используются для косметики (показать мыло, шампунь и т. д.)
Запах имеет яркий аромат. Ребята вам нравится аромат? Что про него можно сказать, какой он (сильный, яркий, приятный, резкий) .
Ребята, а давайте подберем красивые слова для описания ель? (зеленая, хвойная, пушистая, стройная, высокая) . А как мы можем описать сосну ? (высокая, одинокая, могучая, смолистая) .
Правильно, молодцы. А теперь я предлагаю сесть на стульчики.
Воспитатель : Вика прочтет стихотворение Ирины Токмаковой о сосне :
Сосны до неба хотят дорасти,
Небо ветвями хотят подмести,
Чтобы в течение года
Ясной стояла погода.
Воспитатель : И правда сосны бывают очень высокими и это можно увидеть на картине великого художника Ивана Ивановича Шишкина «Корабельная роща» . А знаете, почему эту картину так назвали?
Дети : Нет
Воспитатель : Потому, что из сосен делали мачты для кораблей.
Воспитатель : А про ель стихотворение Гончарова Николая, прочтет Захар
Какой бы лютый холод не был,
И как бы вьюги не гудели, –
Стоят и смотрят гордо в небо
Зеленые, как летом, ели.
Воспитатель : Ребята, а на территории детского сада растут эти деревья? Сколько их на территории садика?
(Ответы детей)
Воспитатель : Давайте вспомним, в чем их сходство?
Ребенок : У сосны и ели одинакового цвета иголки (зеленые, но у сосны светлее , а у елки темнее. Напомните мне, пожалуйста, как можно назвать сосну и ель одним словом ?
Дети : Вечнозеленые.
Воспитатель : Мы много узнали о ели и сосне . И можете их отличить друг от друга. Как вы думаете, удачным ли было наше путешествие? А вы расскажите о нем своим родным? А что вы расскажите? Молодцы!
А теперь я предлагаю отправиться на прогулку и полюбоваться нашими вечнозелеными красавицами.
Как известно, леса – это «легкие планеты». А самая первая ассоциация при слове «лес» – поросль сосны и ели. Без этих деревьев мы слабо представляем свою жизнь. Мы высаживаем их вдоль дорог, чтоб защитить себя от действия угарных газов, делаем мебель, используем при строительстве. В хвойных лесах содержится самое большое разнообразие грибов. Масло хвойных деревьев используется при лечении огромного количества заболеваний. В чем же главные отличия ели от сосны?
Существует большое разнообразие пород, как ели, так и сосны, схожих друг с другом и совершенно различных. Но есть особенности, которые присущи только тому или иному дереву не зависимо от сорта, условий произрастания, возраста. В чем же разница между сосной и елью?
Сосна
Сосна – относится к семейству Сосновых, Класса Хвойные. Распространена в холодном и влажном климате северного полушария, образуя обширные массивы леса. Это хвойное вечнозеленое дерево живет 300-350 лет и за это время достигает 75 метров в высоту. Однако среди сосен встречаются и долгожители. В США, на юго-западе страны, растет сосна, возраст которой насчитывает около 6 тысяч лет.
Сосна совершенно нетребовательна к почве. Может расти как на болотах, так и на сухих песках, на меловых склонах, гранитных скалах, над оврагами. Она легко переносит переизбыток влаги, и растет на самых бедных, песчаных и каменистых грунтах с низким содержанием гумуса. Растение не боится заморозков, засухи, устойчива к ветрам и ураганам. Главное требование дерева к условиям содержания – наличие достаточного количества солнечных лучей. Растение не выносит даже легкого затемнения.
Сосна характеризуется мощной стержневой корневой системой с одним головным корнем, уходящим глубоко в землю. От него расходится огромное количество боковых корней. Такое строение корневой системы сосны помогает сосне доставать воду с больших глубин и осваивать неподходящие для других растений участки земной поверхности.
Сосна обыкновенная
Хвоинки сосны длинные и узкие, располагаются парами. Также, парами, они и опадают. Самое массовое сбрасывание хвоинок происходит осенью. На ветках хвоинки держатся 2-3 года. Для дерева характерно мутовчатое расположение веток – около 5-ти скелетных веток, которые веерообразно расходятся от ствола на одном уровне.
У сосны шишки встречаются двух видов: женские и мужские. Весной на молодых побегах можно наблюдать светло-желтое соцветие мужских шишечек. Они не большие, размером с вишневую косточку. Женские шишечки имеют размер чуть больше булавочной головки и располагаются на конце лапки по одной. Их достаточно сложно различить среди хвоинок на ветке.
Ель
Ель принадлежит к семейству Сосновых, классу Хвойные. Это — вечнозеленое высокое дерево с пушистой пирамидальной кроной. Ели широко распространены в Восточной Европе, Центральной и Северо-Восточной Азии, Северной Америке и Китае. Кроме того, ель является одной из лесообразующих пород. Живет ель в среднем 300 лет и вырастает до 50 метров.
Расположение веток у ели мутовчатое. Каждый год появляется очередная мутовка. К скелетным ветвям крепятся игловидные, четырехгранные хвоинки темно-зеленого цвета. Длина одной хвоинки не превышает 3-х см. Она может крепиться к ветке сроком до 12 лет. Еловая хвоя опадает и заменяется постепенно. У дерева нет выраженного листопадного периода.
Ель
Ель теневынослива, поэтому может расти в смешанном лесу, в тени других деревьев. Дерево может проживать на бедных на питательные вещества каменистых, известковых и подзолистых почвах. В требованиях к влажности почвы это дерево не переносит крайностей. Ни избыточно увлажненный грунт, ни пересушенный, ему не подходит.
Корневая система ели относится к стержневому типу. Но главный корень у дерева развит слабо, а после десяти лет начинает отмирать. Поэтому основной удар на удержание растения в грунте и добывание из него воды и минеральных соединений приходится на боковые корни. С первой задачей корни справляются не всегда, поэтому случаи, когда ель при шквальном ветре выворачивает « с корнем» не одиноки.
У ели шишки встречаются двух видов: женские и мужские. Женские шишки ели сразу бросаются в глаза своим ярко-красным окрасом и размером с лесной орех. Располагаются в верхней части кроны на самых кончиках ветвей. Мужские шишки мельче, их окрас не столь яркий, в них созревает желтая пыльца.
Выводы сайт
- Сосна и ель принадлежат к разным родам одного семейства Сосновые.
- Сосна требует большого количества солнечных лучей, ель – теневыносливое растение.
- Сосна имеет более сильную корневую систему и более устойчива по сравнению с елью.
- Хвоя ели короче и темнее по цвету, чем у сосны.
- У ели нет выраженного листопадного периода, хвоя сосны массово опадает осенью.
Как нарисовать ель
Простое, пошаговое руководство по рисованию елиНажмите ЗДЕСЬ, чтобы сохранить учебник в Pinterest!
Ель – это разновидность вечнозеленого дерева, произрастающего в Северной Америке и Европе. Как и у сосны, у него иглы, а не листья, и он остается зеленым всю зиму. Он также производит семена в свисающих шишках.
Самым старым живым деревом в мире является ель в Швеции, которому около 10 000 лет.
Слово ель происходит от названия региона Пруссия, в которую входили части современной Германии, России и других стран.Область была домом для этого типа дерева. Когда-то все товары, ввозимые из Пруссии, назывались «елью». Вот почему термин «украсить», означающий «сделать новое или красивее», все еще используется сегодня.
Прокрутите вниз, чтобы загрузить этот учебник в формате PDF.
Ель, сосна, кедр, можжевельник, пихта – для некоторых они просто новогодние елки. Когда на выбор так много вечнозеленых деревьев, как вы можете отличить их?
У сосны несколько игл растут из одной почки.У ели и пихты есть индивидуальные потребности в выращивании, но хвоя пихты плоская, тогда как хвоя ели имеет четыре стороны. У можжевельника голубые ягоды, а колючки настоящих кедров расположены по спирали.
Хотите нарисовать величественную ель? Это простое пошаговое руководство по рисованию сделает это проще простого. Все, что вам понадобится, – это ручка или карандаш и лист бумаги. Вы также можете раскрасить законченный рисунок.
Если вам понравился этот урок, см. Также следующие руководства по рисованию: Cartoon Forest, Pinecone и Cartoon Castle.
Пошаговые инструкции по рисованию ели
Рисунок ели – шаг 11. Начните с рисования верхушки или кроны вашего дерева. Сначала нарисуйте перевернутую букву «V», используя две изогнутые линии. Затем соедините линии внизу, используя серию коротких изогнутых линий, которые встречаются в неровных точках. Фигура должна быть примерно треугольной формы.
Рисунок ели – шаг 22. Затем нарисуйте мутовку или округлый набор ветвей под кроной. Протяните изогнутую линию вниз с каждой стороны короны.Затем соедините линии внизу, используя серию коротких изогнутых линий, которые встречаются в неровных точках.
Рисунок ели – шаг 33. Нарисуйте еще два мутовки, каждый немного больше предыдущего. Для каждого оборота вытяните изогнутую линию вниз с каждой стороны предыдущего оборота. Затем соедините линии внизу, используя серию коротких изогнутых линий, которые встречаются в неровных точках.
Рисунок ели – шаг 44. Нарисуйте большую мутовку под предыдущей. Вытяните изогнутую линию вниз с каждой стороны предыдущего оборота, затем соедините линии внизу, используя серию коротких изогнутых линий, которые пересекаются в неровных точках.
Рисование ели – шаг 55. Нарисуйте еще один виток, используя две изогнутые линии, соединенные серией коротких изогнутых линий, которые пересекаются в неровных точках. Обратите внимание, что мутовки больше не становятся больше, чем предыдущие.
Рисунок ели – шаг 66. Нарисуйте еще один оборот, немного меньшего размера, чем тот, что над ним.
Рисунок ели – шаг 77. Нарисуйте еще одну небольшую завитушку под предыдущей частью.
Рисование ели – шаг 88. Нарисуйте еще два завитка, возвращаясь к обычаю делать их больше, чем предыдущие.Обратите внимание, что средняя часть последнего оборота выше, чем по краям. Это позволит создать ствол дерева.
Рисунок ели – шаг 99. Нарисуйте ствол ели двумя изогнутыми линиями. Обозначьте развилки в ветвях V-образной линией с закругленным концом. Затем нарисуйте землю под деревом. Используйте перекрывающиеся изогнутые линии, дополненные фрагментами более коротких линий, пересекающихся в неровных точках. Они указывают на пучки травы.
Полный чертеж ели10. Раскрасьте ель.Ели часто имеют темно-зеленый или даже сине-зеленый цвет.
Хотите нарисовать больше деревьев или даже лес, населенный дружелюбными лесными существами? Ознакомьтесь с нашей подборкой руководств по рисованию растений и животных, чтобы узнать больше.
Прокрутите вниз, чтобы загрузить этот учебник в формате PDF.
Учебное пособие по рисованию для печати
УСТРАНЕНИЕ НЕПОЛАДОК УЧАСТНИКА
Все еще видите рекламу или не можете загрузить PDF-файл?
Сначала убедитесь, что вы вошли в систему.Вы можете войти в систему на странице входа в систему.
Если вы по-прежнему не можете загрузить PDF-файл, вероятным решением будет перезагрузка страницы.
Это можно сделать, нажав кнопку перезагрузки браузера.
Это значок в виде круглой стрелки в верхней части окна браузера, обычно в верхнем левом углу (вы также можете использовать сочетания клавиш: Ctrl + R на ПК и Command + R на Mac).
3.2.A2 Создайте реалистичную ель
Ресурс:
Модуль 3.1 Введение в шейдинг
Расходные материалы:
бумага, карандаши 2H, HB, 2B, 4B и 6B, точилка для карандашей, блок наждачной бумаги, винил и ластики с замесом
ArtSpeak
Ракурс: Компонент перспективы, описывающий искажение объектов и форм жизни при просмотре с экстремальных углов.
Термин «ракурс» применяется к отдельному объекту или фигуре, тогда как термин «перспектива» относится ко всей сцене.
Ель бывает разных форм и размеров (рис. 1). Несмотря на то, что их пропорции разные, все они выглядят как ели, поэтому вы можете нарисовать дерево любого размера и формы.
Рисунок 1
1. Карандашом HB слегка нарисуйте линию в виде центра ствола дерева от низа области рисования почти до вершины.
Рамка вокруг рисунка 2 показывает прямоугольную форму области рисования, используемой для рисования дерева в этом руководстве.
Не пытайтесь провести линию идеально прямой; деревья выглядят более естественно с несколькими изгибами и закруглениями стволов.
Рисунок 2
2. Используйте сквирклы, чтобы добавить землю или основу, на которой растет дерево.
Вы можете сделать землю неровной или достаточно ровной, варьируя форму и размер сквиркул.
3. Используйте сквирклы, чтобы постепенно увеличивать ширину ствола дерева сверху вниз (рис. 3).
Ствол очень тонкий вверху и широкий внизу.
Рисунок 3
4. Добавьте несколько крошечных веток на верхушку ствола дерева.
См. Рис. 5 и увеличенное изображение на рис. 6. Чем дальше от ствола, тем больше каждая ветвь сужается.
Помните старое выражение: «меньше значит больше»! Позже легко добавить новые ветви, если ваше дерево выглядит слишком редким, но стереть слишком полные или толстые ветви очень сложно.
В стороне
Деревья с тонкими листьями (иголками) и шишками называются хвойными.Традиционные рождественские елки в холодном климате обычно хвойные, потому что они остаются зелеными круглый год (вечнозеленые).
Сосна, пихта, можжевельник, лиственница, ель и тис относятся к семейству хвойных.
Во время рисования постоянно напоминайте себе, что ветви ели также растут наружу от ствола спереди и сзади, а не только по бокам.
Например, глядя на дерево с одной точки обзора, вы не можете сказать, насколько широки или длинны ближайшие к вам ветви.
Элемент перспективы, называемый ракурсом, создает иллюзию того, что ветви, которые растут к вам, намного короче, чем они есть на самом деле.
Кроме того, ветви сзади растут от вас и поэтому кажутся меньше, чем ветви спереди и по бокам.
Эти задние ветви также частично скрыты от глаз другими ветвями и стволом дерева.
Рисунок 4
Рисунок 5
5. Добавьте дополнительные ветви к средней части ствола (рис. 6–9).
Не забудьте добавить ветви, которые кажутся перед стволом и позади него.
Рисунок 6
Рисунок 7
Совет!
Чтобы ель выглядела более реалистично, нарисуйте нижние ветви шире и длиннее, чем высокие.
Совет!
Держите под рукой точилку для карандашей и блок наждачной бумаги, чтобы кончики карандашей оставались острыми.
Рисунок 8
Рисунок 9
В стороне
Много раз попрактиковавшись в сквирлинге и изучении елей в природе, вы можете легко научиться рисовать дерево за пару минут.
Тем не менее, вы должны продвигаться к выполнению этого упражнения медленно, чтобы дать себе возможность полностью понять процесс.
6. Добавьте большие ветви к нижней части дерева (рис. 10 и 11).
Пара самых нижних ветвей кажется короче, чем те, что над ними. Эти ветви находятся либо перед стволом, либо за ним.
Рисунок 10
Рисунок 11
7. Добавьте несколько волнистых и рваных линий, идущих от земли, чтобы они выглядели как кусты и трава (рис. 12 и 13).
Рисунок 12
Рисунок 13
Вызов!
Изучите свой рисунок и представьте, как ваше дерево растет в поле. Поздний день, и солнце светит справа, создавая темные тени на некоторых участках дерева.
Солнце начинает садиться, но все еще достаточно высоко, чтобы его свет не мог достигать нижних сторон ветвей.
Солнечные лучи просвечивают сквозь ветви, и некоторые участки ствола справа кажутся светлее.
Левая часть ствола в основном в тени.
Отбрасываемая тень от дерева затемняет листву слева и под деревом. Трава и кусты справа (ближе к солнечному свету) светлее, чем в тени.
8. Используйте карандаши 4B и 6B и сквирклыки, чтобы добавить тени к некоторым участкам ветвей, ствола и наземной листвы (рис. 14–17).
Не добавляйте дополнительное затемнение к светлым участкам, расположенным ближе к солнцу.
Также будьте осторожны, не добавляйте слишком много теней!
Темное затенение следует добавлять умеренно – только в теневые участки.
Рисунок 14
Рисунок 15
Рисунок 16
Рисунок 17
Подпишите свой рисунок и поставьте дату, крепко обнимите себя и идите обнимать дерево.
Совет!
Всякий раз, когда у вас есть возможность посетить лесную местность, найдите время, чтобы обойти отдельные деревья и рассмотреть их ветви с разных точек зрения.
Вызов!
Вынесите альбом и карандаши на улицу ближе к вечеру в солнечный день.
Найдите дерево и нарисуйте его десять раз.
Меняйте свое положение каждый раз, когда вы начинаете новый рисунок, чтобы солнечный свет выглядел падающим с разных сторон.
Нарисуйте свет и тени в точности такими, какими вы их видите.
Хвойные от художника-пейзажиста Клаудии Ниццы
Автор и художник Клаудиа Найс (www.brightwoodstudio.com) опубликовал множество учебных пособий по искусству. Следующий отрывок появился в журнале The Artist’s Magazine (сентябрь 2011 г.) и взят из книги Рисование и рисование деревьев в пейзаже . Прокрутите вниз, чтобы узнать ее советы по рисованию хвойных деревьев, а также ознакомиться с ее тремя новыми DVD-дисками для художников (включая основы рисования текстур). ~ Чери
Убедительные хвойные деревья: как рисовать деревья Клаудиа НиццаПоскольку большинство рисунков, выполненных карандашом и пером, выполнены в серых и черных тонах, наиболее важными факторами для создания хорошей репрезентации являются линии и тональный контраст.Отметка или линия, нанесенная инструментом рисования, отвечает за определение края и общей формы дерева, его шероховатости или гладкости, а также его положения или движения. Насколько темная метка и насколько близко нарисованы линии, отвечает за создание контраста по значению – относительные серые и черные тона на рисунке. Это значение контраста, которое придает рисунку трехмерный вид, описывая моделирующие тени дерева, отбрасываемые тени и текстуру поверхности.
Иногда называемые хвойными или вечнозелеными деревьями, хвойные деревья – это деревья с иголками или чешуевидными листьями.Хотя у большинства из них листья сохраняются круглый год, некоторые, например лиственница и тамарак, становятся золотыми и опадают хвоей поздней осенью. Лиственная часть (крона) хвойных обычно несколько конусовидная. Однако угол и размах конечностей, а также форма туловища могут сильно различаться от вида к виду. Линии каракулей хорошо сочетаются с текстурой деревьев на заднем плане.
Прежде чем рисовать карандашом на бумаге, начните с изучения своего дерева.Какой он рост? Какова форма его внешнего периметра? Ветви достигают неба, тянутся горизонтально или опускаются к земле? Ствол прямой или изогнутый? Изучите молодую калифорнийскую красную ель на фотографии (ниже). Затем выполните следующие действия, чтобы нарисовать простое хвойное дерево.
1. Карандаш в багажнике. Начните с легкого карандаша по высоте и форме туловища. Это саженец слегка изгибается вправо.
2. Нарисуйте корону. Слегка нанесите карандашом общую форму кроны дерева. Обратите внимание, как далеко он простирается вниз по стволу. Карандашом отметьте основные ветки. Обратите внимание, что не все ветви доходят до внешнего края кроны. Ветви, идущие вперед, обозначены линией (A), проходящей через ствол.
3. Залейте листву. Задрапируйте ветки пятнами листвы, отмечая тени немного сильнее. Во время работы стирайте направляющие ствола и ответвления, кроме тех мест, где они пересекают открытые участки.Для более мягкого вида используйте бумажный пень или лепешку, чтобы растушевать участки листвы. ~ CN
И вот отличные новости – Клаудиа Найс только что выпустила три новых DVD! Щелкните следующие ссылки, чтобы узнать, что подходит именно вам.
С уважением,
Cherie
Научитесь рисовать сосну
Рисунок сосны – Pinus silvestris
Десять характерных черт сосен объясняются и показаны на большом изображении эскиза сосны.В уроке объясняется, как нарисовать сосну с верхушки дерева иголками, ветви к стволу с разными зонами коры. Примерный рисунок был выполнен пером на месте прямо с натуры. Узнайте, как нарисовать реалистично выглядящую сосну. Посмотреть больше примеров, почерпнуть вдохновение для собственных рисунков можно здесь.
Для тех, кто ищет дешевую книгу по рисованию деревьев, я могу порекомендовать мою публикацию на Blurb «Как нарисовать дерево»
Если вы искали ботаническую информацию, я могу порекомендовать другую линзу Squidoo Сосна обыкновенная
Изображение предоставлено: Все изображения на этой веб-странице, если не указано иное, созданы автором.Изображения и иллюстрации продуктов (в партнерских ссылках) используются согласно Squidoo TOS.
Пример рисунка сосны – объяснение 10 характерных особенностей
Рисование пером
Ниже приведен этюд из альбома для рисования пером, изображающий сосну. Если вы нажмете на изображение, большая его версия с текстом откроется в новой вкладке. Возможно, вы захотите скачать и распечатать этот файл (около 2 МБ) и изучить его. На эскизе отмечены десять цифр с пояснениями, которые указывают на некоторые типичные особенности сосен.Конечно, внешний вид сосен варьируется в зависимости от вида, возраста, окружающей среды и условий роста, но если вы поймете и распознаете важные особенности, будет легко нарисовать свой собственный рисунок.
Пояснения к чертежу сосны с изображениями деталей
1. Я начал рисовать сосну сверху. Сначала была нарисована форма верхушки дерева в виде контура. Я посмотрел на дерево, а не на свою бумагу, и проследил за контуром, который мог видеть на фоне светлого неба.Это называется слепым розыгрышем, если вы не знаете об этом, проверьте ссылку.
2. Затем я попытался определить узор, ритм или структуру иголок сосны и выполнил эти детали в рамках ранее нарисованного контура
3. Я заметил 3-4 основных закономерности во внешнем виде игл и записал их. Вы можете видеть вещи по-другому и можете создавать свои собственные знаки или знаки для представления. Возможно, вы захотите поместить их на отдельный лист.
4.Я узнал, что по краям больших гроздей есть тонкие ветки с иголками и без них. Эти тонкие ветки немного похожи на паутинную паутину и придают типичный вид сосне.
5. Затем я продолжил вниз по контуру ствола и добавил ветви. Ветви сосен делают типичные повороты, другие деревья так не делают. По этой причине сосну можно распознать на много миль.
6. Есть также много старых коротких веток, которые тоже создают типичный вид.
7. Более крупные ветви имеют хорошо заметные «плечи» на стыке со стволом.
8. Каждый год вокруг ствола разрастается несколько ветвей, округляющих его. Ежегодный рост в высоту все еще виден по длине свободного пространства между двумя из этих кругов, что можно увидеть по этим маленьким темным отметкам ветвей на коре, даже когда ветки уже отсутствуют.
Кора сосны меняется сверху вниз. в верхней части он яркий, а иногда и блестящий.Затем он меняется в области красноватой поверхности, где тонкая мягкая кора выступает из кусков старой кожи.
9. Далее кора становится более темной и шероховатой, с множеством мелких морщинок на поверхности.
10. Внизу кора очень толстая, с типичными глубокими трещинами и щелями.
11. Для сравнения я добавил небольшой кусок ели на заднем плане. Наблюдая и изображая типичные черты дерева, можно вызвать реалистичное впечатление, даже если пропорции не совсем правильные.
Сосна на рисунке пейзажа
Это рисунок индийской тушью, сделанный холодным январским утром. Молодая сосна – важная часть декорации. Это другой вид (Pinus niger) с более густой верхушкой дерева. На деталях ниже можно увидеть, как массы игл были представлены простыми штрихами в разных направлениях.
Как нарисовать дерево – Книга опубликована на BLURB
Полевое руководство по рисованию и зарисовке деревьев
Маленькая книжка формата 8X5 основана на содержании этого объектива.Содержит
70 черно-белых изображений рисунков пером, тушью и углем. В книге вы найдете множество иллюстраций с помощью этого объектива, а также другие иллюстрации из связанных объективов по рисованию лиственных и хвойных деревьев, а также некоторые дополнительные изображения.
В книге обобщен мой опыт рисования деревьев. На большинстве рисунков изображены лиственные деревья, но есть также пояснения по поводу сосны и ели.
Ссылка на книгу Как нарисовать дерево Доступен полный превью.
Прекрасные образцы рисунков сосны, выполненные художниками на FLICKR
Особый совет !!!!!
Я рада, что случайно нашла эти вдохновляющие рисунки. Я рад, что тем временем автор добавил свое имя. Все эти великолепные рисунки выполнены Майком де Визом, профессиональным иллюстратором.
Я пишу эти абзацы только для того, чтобы убедиться, что эта ссылка не пропущена!
Набор пленэровых рисунков тушью
Этот набор пленэров, опубликованных пользователем FLICKR Майком ДеВизом, содержит серию великолепных рисунков, в основном сосны, но также и других хвойных пород.Художники изучили все части дерева от ствола до верхушки дерева, включая детали и целые леса.
Очень вдохновляющий набор работ!
Как нарисовать сосну – видео инструкция
Видео для начинающих – рисование сосны тушью и кистью
Сосна – сосна от yanghaiying
Чертежи сосны онлайн – список ссылок
Архив рисунков сосны
Сосна – Архив американского искусства: Альбомы для рисования: Выбор кураторов
Альбом для рисования Уортингтона Уиттреджа о путешествии по Рейну 1849 г. / Уортингтон Уиттредж, художник.Альбом для рисования: 1 т .: различные носители; 15 х 24 см. Документы Уортингтона Уиттреджа. Архивы американского искусства.
Исследование сосны на Flickr – обмен фотографиями!
Красивое исследование Art * край на FLICKR
# Мне понравились дикие линии этой сосны. Так полно текстуры. Акварель и тушь.
Рисунок городского пейзажа
Изучение города с альбомом для зарисовок
С апреля 2006 г. по май 2007 г. я работал над выставочным проектом о городском ландшафте Штутгарта, Германия.
Я выбрал более 30 рисунков для книги, которая теперь доступна на сайте BLURB.
На первых 10 страницах показаны панорамные рисунки вместе с эссе о городском пейзаже и опыте рисования такого сложного предмета. На следующих страницах показаны два панорамных рисунка на странице без текста.
Другие линзы для рисования от Edition Handdruck
Пейзажный рисунок – улучшайте свои навыки быстро и качественно!
8 советов, как эффективно и быстро улучшить свои навыки рисования пейзажей.Каждый, кто может написать свое имя на листе бумаги, может достичь высокого уровня навыков рисования. Рисовать гораздо проще, чем рисовать, и это доставляет огромное удовольствие! Ссылка на некоторые из моих эскизов и рисунок
Как нарисовать дерево
Как нарисовать дерево – Найдите коллекцию рисунков деревьев и объяснения того, как они были сделаны. Вдохновляйтесь известными примерами и просмотрите онлайн-уроки рисования. Найдите свой способ нарисовать дерево.
Как рисовать лиственные деревья
Это введение в рисование лиственных деревьев с натуры.MPM, художник по гравюрам Monotype Мартин объясняет, как он рисует лиственные деревья ручкой и углем. В рисовании нет правил, каждый может придумать свой способ рисования дерева. Если вы ищете вдохновения или идеи, возьмите туалет
Альбомы для рисования – мелочи превращаются в большие дела
В течение десятилетий альбомы для рисования художников были замечены только ограниченной специальной аудиторией, которая получила доступ к этим драгоценным ресурсам в основном только после смерти создателя.Выдержки могли быть опубликованы в печатных СМИ, но в основном только ценой повреждения или уничтожения альбомов.
Эскизное снаряжение – что нужно новичку?
Рисование на открытом воздухе – отличное занятие и полезное хобби для всех. Этот объектив для тех, кто хочет рисовать на улице в городе или в сельской местности. Какие инструменты для рисования вам понадобятся? Какое снаряжение подходит и каким должно быть качество?
Размер хвои ели европейской и изменчивость формы поперечного сечения, вызванная облучением в макро- и микромасштабе и концентрацией CO2
Trees (2018) 32: 231–244
DOI 10.1007 / s00468-017-1626-3
ОРИГИНАЛЬНАЯ СТАТЬЯ
Размер хвои ели обыкновенной и изменение формы поперечного сечения
, вызванное излучением в макро- и микромасштабе и концентрацией CO2
Zuzana Kubínová1 · JiříJanáček2 · ZuzanaLhotáková1 · MiroslavaŠprtová3 ·
LucieKubínová2 · JanaAlbrechtová1,4
Поступила в редакцию: 20 октября 2017 г. Октябрь 2017 г.
© Springer-Verlag GmbH, Германия, 2017 г.
Иглына макроградиенте освещенности были плоскими
и имели меньшую площадь поперечного сечения и меньший объем, чем иглы
для солнечных лучей.Микроградиент освещенности был обнаружен
как на солнечных, так и на теневых побегах, более крутой на солнечных побегах
, чем на теневых побегах. В микромасштабе градиент освещенности
вызвал изменения размера иглы и формы поперечного сечения
в соответствии с ориентацией игл на побеге. Из-за более благоприятной световой среды
черты верхней иглы
как на солнечных, так и на теневых побегах больше походили на солнечные
черты иглы.Объем солнечной иглы был значительно больше
в случае повышенного [CO2] по сравнению с окружающим [CO2].
Облучение было более сильным морфогенным фактором, определяющим размер иглы
и форму поперечного сечения по сравнению с концентрацией CO2
. Мы продемонстрировали, что обобщенный анализ Procrustes
может быть очень мощным инструментом в экофизиологических исследованиях
для оценки мелкомасштабных, едва различимых изменений формы листьев
на внутривидовом уровне, вызванных факторами окружающей среды.
Ключевые слова Морфология хвои · Повышенная
концентрация CO2 · Обобщенный анализ Прокруста ·
Геометрическая морфометрия · Градиент освещенности · Форма листа
Введение
уменьшает освещенность, тем самым влияя на градиент освещенности
и, таким образом, развитие солнца и тени оставляет
(Шталь 1883; Ким и др. 2005). У видов с дорсивентральными листьями
солнечные листья более толстые по сравнению с теневыми листьями, при этом большая толщина на
сопровождается большим количеством слоев мезофилла pali-
sade (Wylie 1949; Hanba etal.2002; Kim etal.
2005; Нинемец 2007) и большей внутренней поверхности мезофилла
площади (Терашима 2005). Некоторые виды хвойных также демонстрируют не-
различий между солнечными и теневыми экотипами, но
Abstract
Ключевое сообщение Параметры поперечного сечения иглы отличаются на
в зависимости от ориентации иглы на побеге, соответствующей микроградиенту освещенности. Излучение
является более сильным морфогенным фактором, определяющим размер и форму иглы
, чем концентрация CO2.
Резюме Мы исследовали влияние облучения на макро- и микромасштабные мак-
и микромасштабы, повышенную концентрацию СО2 [СО2],
и их взаимодействие на хвое ели европейской (Picea abies L.
Karst.). Макромасштаб освещенности
был представлен солнечными и теневыми побегами из двух вертикальных положений в коронке
, а микромасштаб освещенности соответствовал пространственной ориентации отдельных игл на побеге
(верхняя, боковая,
и нижняя иглы относительно иглы
). ось выстрела).Определение
формы поперечного сечения иглы с использованием обобщенного анализа Procrustes
и анализа главных компонентов обеспечило новый подход
к оценке морфометрии иглы. Как и ожидалось,
Сообщил Г. Пиовесан.
Электронные дополнительные материалы Онлайн-версия этой статьи
(doi: 10.1007 / s00468-017-1626-3) содержит дополнительные материалы
, которые доступны авторизованным пользователям.
* Яна Альбрехтова
albrecht @ natur.cuni.cz
1 Кафедра экспериментальной биологии растений, факультет
науки, Карлов университет, Viničná 5, 12844Prague2,
Чешская Республика
2 Институт физиологии Чешской академии ofSciences,
vvi, Vídeská 1083, 14220Prague4, CzechRepublic
3 Институт исследований глобальных изменений, Чешская академия
ofSciences, vvi, Bělidla 986 / 4a, 60300 Брно,
Чешская Республика
4 Институт Ботаники Чешской академии наук, т.v.i.,
Zámek 1, 25243Prhonice, CzechRepublic
Содержание предоставлено Springer Nature, применяются условия использования. Права защищены.
Анатомия и морфология развития вегетативных почек на отдельных ветвях ели европейской в контролируемых условиях до распускания почек | Физиология деревьев
Абстрактные
Мы исследовали световую и стереомикроскопическую структуру развивающихся вегетативных почек у 16-летней ели европейской [ Picea abies (L.) Karst.] Южно-финского происхождения в связи с суммой температур и внешне видимыми изменениями почек до и во время распускания почек в условиях выгонки. Ветки были собраны 17 января и перенесены в теплицу, где они были сначала подвергнуты предварительным условиям (темнота, +4 ° C) в течение 7 дней, а затем – условиям выгонки (продолжительность дня 12 ч, +20 ° C). В период с 17 января по 13 февраля образцы почек собирали 20 раз. Температура воздуха регистрировалась ежечасно на протяжении всего периода исследования.Первым микроскопическим изменением было временное увеличение размера и количества липидных капель перед началом накопления температурной суммы ( T ≥ +5 ° C). С 4-го по 9-й день в условиях выгонки трахеиды начали развиваться от основания до верхушки почки. Это было тесно синхронизировано с наблюдаемым морфологическим изменением формы кончика иглы с закругленной на заостренную. Развитие от первого видимого изменения чешуи почек на 12-й день выгонки до распускания почек длилось 9 дней при сумме температур 313 дней.d. Суммы температур в нашем эксперименте завышали требования к сумме температур для фаз развития бутонов, измеренные в поле. Развитие бутона можно разделить на четыре структурных этапа. Первые две фазы, т. Е. Морфологические изменения первичной хвои, произошли без каких-либо внешне видимых изменений почек. Таким образом, эти фазы имеют потенциал для тестирования и улучшения фенологических моделей, которые до сих пор в основном основывались на наблюдении за распусканием почек невооруженным глазом.
Введение
Деревья, растущие в северной бореальной зоне, сталкиваются с сезонными колебаниями температуры и освещенности, поэтому период активного роста и морозостойкое состояние покоя чередуются в их годовом цикле (Weiser 1970, Sarvas 1972, 1974, Fuchigami et al. 1982). Правильный выбор времени перехода от восприимчивого состояния активного роста к морозостойкому состоянию покоя и обратно имеет фундаментальное значение для выживания деревьев и регулируется взаимодействием между генетическими факторами и факторами окружающей среды.
После прекращения роста осенью вегетативные почки деревьев переходят в состояние покоя, которое обычно делится на три последовательные фазы (Lang et al. 1987). Осенью и зимой характер и глубина покоя постепенно меняются. Осенью во время развития покоя почки входят в состояние эндодормирования, в течение которого они неспособны к росту, даже когда благоприятные условия для выращивания временно возвращаются (Doorenbos 1953, Samish 1954, Lang et al. 1987). Освобождение от эндодонтии, которое требует длительного воздействия низких температур охлаждения (например,g., от -5 до +10 ° C), за ним следует экологический запах, который характеризуется физиологическими изменениями и внутренней способностью расти и развиваться в направлении распускания почек (Romberger 1963, Weiser 1970, Kobayashi and Fuchigami 1983, Hänninen 1990) . Экологический запах поддерживается неблагоприятными низкими температурами окружающей среды и высвобождается за счет воздействия высоких температур (например, T > +5 ° C) (Smith and Kefford 1964, Perry 1971, Fuchigami et al. 1982). Распространение бутонов происходит, когда сумма накопленных температур превышает пороговое значение для конкретного вида (Sarvas 1972, Campbell and Sugano 1975, Cannell and Smith 1983).Таким образом, начало роста весной в основном регулируется температурой воздуха (Sarvas 1972), хотя световые условия, такие как продолжительность дня, также играют роль (Heide 1993 a , 1993 b , Häkkinen et al. 1998, Partanen и др., 1998, 2001).
Время выхода из эндодормальности почек и перехода к экодормальности, а также снижение морозостойкости во время развития почек имеют решающее значение для деревьев, произрастающих в северных и умеренных регионах (Sarvas 1972, 1974, Fuchigami et al.1982, Kalberer et al. 2006 г.). Слишком раннее начало роста и обезвоживание весной может вызвать повреждение морозом и плохую выживаемость, тогда как отсроченное начало роста приводит к неэффективному использованию вегетационного периода и к плохой конкурентоспособности (Heide 1985). Прогнозируемое глобальное потепление в средних и высоких широтах, особенно в зимние месяцы (Болин и др., 1986, Шнайдер, 1989, Картер и др., 1995), может сделать бореальные леса более восприимчивыми к морозам из-за учащения случаев колебания температуры ранней весной (Ahas et al.2002). Это может нейтрализовать более положительные эффекты повышения продуктивности, связанные с более теплым и продолжительным вегетационным периодом (Jarvis and Linder 2000). С другой стороны, было показано, что относительно высокие осенние температуры задерживают распускание почек следующей весной у бореальных широколиственных пород деревьев (Heide 2003).
Время распускания почек в зависимости от нынешних и будущих температур сообщалось в нескольких исследованиях как на хвойных, так и на лиственных деревьях. Однако большинство из них основано на видимых фенологических изменениях в бутоне, т.е.g., набухание почек, распускание почек и распускание листьев (например, Linkosalo et al. 2000, Slaney et al. 2007). Модели развития бутонов использовались в различных типах моделирования для прогнозирования возможных последствий глобального потепления климата (Hänninen 1991, 1995, Kellomäki et al. 1995, Linkosalo et al. 2000). В большинстве моделей начало развития вегетативных почек во время экодормирования основывается не на наблюдениях, сделанных внутри почек, а на теоретических допущениях или оценках, экстраполированных на основе наблюдений за распусканием почек (Cannell and Smith 1983, Häkkinen et al.1998). Чтобы оценить и повысить точность таких моделей, необходимо наблюдать за изменениями, происходящими внутри бутона, прежде чем можно будет увидеть какие-либо внешне видимые признаки.
Изменения вегетативных почек хвойных деревьев от зимы к весне и лету были задокументированы на молекулярном, генетическом, анатомическом, тонком структурном и биохимическом уровнях (Купила-Ахвенниеми и др. 1987, Хейнович и Обарска 1995, Билкова и др. 1999, de Faÿ et al.2000, Svobodová et al.2000, Guzicka and Woźny 2003, Jordy 2004, Gyllenstrand et al.2007, Яковлев и др. 2008 г.). Однако, насколько нам известно, анатомические и морфологические фазы развития от раннего экологического наступления до распускания почек в вегетативных почках ели европейской [ Picea abies (L.) Karst.] Не были подробно описаны на основе ежедневный отбор проб в зависимости от суммарной температуры и внешнего вида бутонов.
Цели данного исследования заключались в (i) выявлении начальных анатомических и морфологических изменений, указывающих на начало развития почек у ели европейской, (ii) описании структурных фаз развития первичных побегов вегетативных почек ели в зависимости от сумма температуры и внешне видимые изменения в почек во время экологического запаха и (iii) для определения анатомических и / или морфологических фаз, которые могут быть использованы в практических полевых исследованиях, в дополнение к визуальному наблюдению за распусканием почек.Мы проследили суточное развитие почек невооруженным глазом, а также с помощью стерео- и светового микроскопа на свежесрезанных и химически закрепленных почках, которые были отделены от ветвей в условиях экспериментальной выгонки.
Материалы и методы
Образец дерева
Ветви были собраны с одного случайно выбранного 16-летнего дерева европейской ели. Дерево растет в испытании потомства (номер 1416/3), установленном на участке типа Myrtillus (Cajander 1949) по соседству с исследовательским центром Пункахарью (61 ° 48 ′ с.ш. и 29 ° 20 ′ в.д.) финской Лесной научно-исследовательский институт.Дерево имеет южно-финское происхождение и выращено из семян, полученных из семенного сада номер 73 в Таммеле (60 ° 39 ′ с.ш. и 24 ° 02 ′ в.д.) и собранных из прививок материнского дерева E 170 из Руоколахти (61 ° 20 ′ Северной широты и 29 ° 00 ′ восточной долготы). Саженец был выращен в питомнике Патама в Саариярви (62 ° 45 ‘с.ш. и 25 ° 38’ в.д.) перед пересадкой в Пункахарью в июне 1993 года. Температуру воздуха регистрировали один раз в час в поле на высоте 2 м над уровнем моря. на уровне земли с регистраторами данных Tinytalk. На рисунке 1 показаны среднесуточная температура воздуха и накопленные холодильные агрегаты (гл.u. h −1 ) (Sarvas 1974) с 1 сентября 2006 г. до начала эксперимента 17 января 2007 г.
Рисунок 1.
Среднесуточная температура воздуха (колеблющаяся линия) и накопленные холодильные агрегаты (восходящая линия) в Пункахарью, Финляндия (61 ° 48 ′ с.ш. и 29 ° 20 ′ в.д.), с 1 сентября 2006 г. до начала эксперимента. 17 января 2007 г. Пунктирная линия обозначает температуру 0 ° C.
Рис. 1.
Среднесуточная температура воздуха (колеблющаяся линия) и накопленные холодильные агрегаты (восходящая линия) в Пункахарью, Финляндия (61 ° 48 ′ северной широты и 29 ° 20 ′ восточной долготы), с 1 сентября 2006 г. эксперимент 17 января 2007 г.Пунктирная линия показывает температуру 0 ° C.
Предварительные и форсирующие условия
17 января 2007 г. 25 ветвей были отделены от второго, третьего и четвертого верхних оборотов ветвей и доставлены в лабораторию исследовательского центра Пункахарью, где их содержали в течение 1 недели в темноте при +4 ° C с помещенными срезанными концами. в воде. После этого периода предварительной форсировки их перевели в условия выгонки в теплицу, где температура воздуха поддерживалась постоянной на уровне +20 ° C, а продолжительность светового дня – 12 часов.Относительно высокая температура выгонки, +20 ° C, использовалась для успешного завершения развития бутонов на отделившихся ветвях. Фотопериод контролировался с помощью черных штор. Для увеличения естественной продолжительности светового дня использовали металлогалогенные лампы (Philips HPI-T Powertone 400 Вт). Фотосинтетически активное излучение на уровне ветвей составляло около 50 мкЭ · м −2 с −1 . Шторы были открыты, а лампы горели с 6 до 18 часов (стандартное время). Чтобы гарантировать подачу воды к веткам, их концы были обрезаны, а воду в ведрах меняли раз в неделю.Температура воздуха регистрировалась один раз в час в условиях предварительного форсирования и форсирования на уровне ответвлений с помощью регистраторов данных Tinytalk.
Отбор и анализ почек
В каждую из 20 дат отбора проб (17 января – 13 февраля) случайным образом отбирали одну ветвь, отрезали пять верхушек побегов длиной около 2,5 см с конечной почкой и окружающими ее почками (бутонами мутовки) и помещали в пробирку. содержащий фиксирующий раствор (2% глутарового альдегида в какодилатном буфере; pH 7.0). Молярность какодилатного буфера доводили до 0,1 М в предварительных условиях, до 0,07 М в течение первых семи дней в условиях воздействия и до 0,05 М до конца отбора проб (Сойккели, 1980). Отбор проб проводился в утренние часы (с 8 до 9 часов утра), за исключением первого образца, который был взят в 13 часов 17 января, когда ветви были принесены с поля. Второй образец был взят 22 января, когда завершился пятый день предварительной форсировки, а третий образец был взят 24 января, когда завершился 7-й день предварительной форсировки, т.е.е., непосредственно перед переводом веток в условия выгонки. Таким образом, первым днем выгонки было 25 января, когда ветви находились в условиях выгонки первые 24 часа. Образец отбирали ежедневно с первого по 15-й день нагнетания, а последние два образца – на 19-й и 20-й дни нагнетания (Таблица 1). Пробирки с образцами были перевезены в сумке-холодильнике в исследовательский отдел Йоэнсуу Финского научно-исследовательского института леса для морфологических и анатомических исследований.
Таблица 1.Анатомо-морфологическая характеристика развивающихся вегетативных почек ели европейской. Наличие липидных капель в клетках (- = событие еще нет / больше нет, + = умеренное и ++ = обильное), развитие трахеид от основания почки до всей почки, смена примордиальных игл на заостренные у основания и вершина почки, видимость верхушки почки, связанная с ростом первичных игл (полностью видимая = нет роста игл, частично видна = рост игл начался и вершина видна только частично, не видна = верхушка полностью покрыта растущими первичными иглами), среднее длина (мм) первичного побега и всей почки и их соотношение.Ветки были собраны и переведены с поля в форсинговые условия 17 января и в форсинговые – 24 января. Накопление суммы температуры [сумма градусо-дней (dd) T ≥ +5 ° C, сумма градус-часов (dh) T ≥ +5 ° C и сумма единиц периода (pu h −1 )] рассчитывалась из 17 января до окончания эксперимента 13 февраля.
| Отбор проб | Накопление температуры | Липиды 1 | Трахеиды развиты до зародыша | Первичные иглы заострены | Верхушка почки | Средняя длина | от первичного побега до целого бутона | |||||||||||
| d.d. | д.х. | о.е. h −1 | Основание | Половина | Верх | Основание бутона | Верх бутона | Полностью видно | Частично видно | Не видно | Прим. побег | Целый бутон | ||||||
| В поле | 0,0 | 0,0 | 0,0 | – | – | – | – | – | – 9068 – 9068 | 2.0 | 7,5 | 0,3 | ||||||
| Предварительный день 5 | 0,0 | 0,0 | 168,6 | – | – | – | – | – | – | – | – | 2,0 | 8,0 | 0,3 | ||||
| Предварительный день 7 | 0,0 | 0,0 | 233,2 | + | – | – | – | – | – | – | – | – | – | 3.9 | 12,5 | 0,3 | ||
| Форсирование дней | ||||||||||||||||||
| 1 | 15.9 | 382.6 | 647.7 | + | – | – | – | – 9068 | – | – | 3,9 | 11,4 | 0,3 | |||||
| 2 | 31,8 | 762,6 | 1059,2 | ++ | – | – | – | × | – | – | 2.0 | 8,4 | 0,3 | |||||
| 3 | 47,5 | 1139,2 | 1466,6 | ++ | – | – | – | – | – | – | – | 2,0 | 8,1 | 0,3 | ||||
| 4 | 63,3 | 1518,8 | 1877,4 | + | × | – | – | – | – | – | 90702.6 | 6,6 | 0,3 | |||||
| 5 2,4 | 79,0 | 1895,2 | 2284,4 | – | × | – | × | × | – | 8,7 | 0,3 | |||||||
| 6 | 94,8 | 2274,0 | 2694,4 | – | × | – | – | × | – | × | – 2 .5 | 7,8 | 0,4 | |||||
| 7 | 110,7 | 2656,7 | 3109,2 | × | – | × | – | × | – | × | – | 7,1 | 0,4 | |||||
| 8 | 126,7 | 3039,7 | 3524,4 | × | – | × | × 6 | (8) | 3.4 | 7,5 | 0,5 | |||||||
| 9 | 142,6 | 3421,4 | 3937,6 | × | × | × | × | 2,9 | 8,2 | 0,4 | ||||||||
| 10 | 158,5 | 3803,5 | 4351,6 | × | × | 3.1 | 9,5 | 0,4 | ||||||||||
| 11 | 174,2 | 4180,9 | 4759,9 | × | × | × | 0,5 | |||||||||||
| 12 | 190,1 | 4562,6 | 5173,4 | × | × | .1 | 9,8 | 0,4 | ||||||||||
| 13 | 205,9 | 4940,8 | 5582,7 | × | 0,5 | |||||||||||||
| 14 | 221,8 | 5322,0 | 5995,8 | × | × | 1 | 9,5 | 0,5 | ||||||||||
| 15 | 237,7 | 5704,7 | 6410,6 | × | × | 0,5 | ||||||||||||
| 19 | 297,9 | 7149,8 | 7967.1 | × | × | 7 | 90.310,1 | 0,8 | ||||||||||
| 20 | 312,9 | 7509,5 | 8354,2 | × | × | 0,8 | ||||||||||||
| Количество бутонов на образец | 3 | 3 | 10 | 10 | 2 | 2 | ||||||||||||
| Отбор образцов | Первичные иглы заострены на | Верхушка бутона | Средняя длина (мм) | Среднее отношение первичного побега к целому бутону | ||||||||||||||
| d.d. | д.х. | о.е. h −1 | Основание | Половина | Верх | Основание бутона | Верх бутона | Полностью видно | Частично видно | Не видно | Прим. побег | Целый бутон | ||||||
| В поле | 0,0 | 0,0 | 0,0 | – | – | – | – | – | – 9068 – 9068 | 2.0 | 7,5 | 0,3 | ||||||
| Предварительный день 5 | 0,0 | 0,0 | 168,6 | – | – | – | – | – | – | – | – | 2,0 | 8,0 | 0,3 | ||||
| Предварительный день 7 | 0,0 | 0,0 | 233,2 | + | – | – | – | – | – | – | – | – | – | 3.9 | 12,5 | 0,3 | ||
| Форсирование дней | ||||||||||||||||||
| 1 | 15.9 | 382.6 | 647.7 | + | – | – | – | – 9068 | – | – | 3,9 | 11,4 | 0,3 | |||||
| 2 | 31,8 | 762,6 | 1059,2 | ++ | – | – | – | × | – | – | 2.0 | 8,4 | 0,3 | |||||
| 3 | 47,5 | 1139,2 | 1466,6 | ++ | – | – | – | – | – | – | – | 2,0 | 8,1 | 0,3 | ||||
| 4 | 63,3 | 1518,8 | 1877,4 | + | × | – | – | – | – | – | 90702.6 | 6,6 | 0,3 | |||||
| 5 2,4 | 79,0 | 1895,2 | 2284,4 | – | × | – | × | × | – | 8,7 | 0,3 | |||||||
| 6 | 94,8 | 2274,0 | 2694,4 | – | × | – | – | × | – | × | – 2 .5 | 7,8 | 0,4 | |||||
| 7 | 110,7 | 2656,7 | 3109,2 | × | – | × | – | × | – | × | – | 7,1 | 0,4 | |||||
| 8 | 126,7 | 3039,7 | 3524,4 | × | – | × | × 6 | (8) | 3.4 | 7,5 | 0,5 | |||||||
| 9 | 142,6 | 3421,4 | 3937,6 | × | × | × | × | 2,9 | 8,2 | 0,4 | ||||||||
| 10 | 158,5 | 3803,5 | 4351,6 | × | × | 3.1 | 9,5 | 0,4 | ||||||||||
| 11 | 174,2 | 4180,9 | 4759,9 | × | × | × | 0,5 | |||||||||||
| 12 | 190,1 | 4562,6 | 5173,4 | × | × | .1 | 9,8 | 0,4 | ||||||||||
| 13 | 205,9 | 4940,8 | 5582,7 | × | 0,5 | |||||||||||||
| 14 | 221,8 | 5322,0 | 5995,8 | × | × | 1 | 9,5 | 0,5 | ||||||||||
| 15 | 237,7 | 5704,7 | 6410,6 | × | × | 0,5 | ||||||||||||
| 19 | 297,9 | 7149,8 | 7967.1 | × | × | 7 | 90.310,1 | 0,8 | ||||||||||
| 20 | 312,9 | 7509,5 | 8354,2 | × | × | 0,8 | ||||||||||||
| Количество бутонов на образец | 3 | 3 | 10 | 10 | 2 | 2 | ||||||||||||
Анатомо-морфологическая характеристика развивающихся вегетативных почек ели европейской. Наличие липидных капель в клетках (- = событие еще нет / больше нет, + = умеренное и ++ = обильное), развитие трахеид от основания почки до всей почки, смена примордиальных игл на заостренные у основания и вершина почки, видимость верхушки почки, связанная с ростом первичных игл (полностью видимая = нет роста игл, частично видна = рост игл начался и вершина видна только частично, не видна = верхушка полностью покрыта растущими первичными иглами), среднее длина (мм) первичного побега и всей почки и их соотношение.Ветки были собраны и переведены с поля в форсинговые условия 17 января и в форсинговые – 24 января. Накопление суммы температуры [сумма градусо-дней (dd) T ≥ +5 ° C, сумма градус-часов (dh) T ≥ +5 ° C и сумма единиц периода (pu h −1 )] рассчитывалась из 17 января до окончания эксперимента 13 февраля.
| Отбор проб | Накопление температуры | Липиды 1 | Трахеиды развиты до зародыша | Первичные иглы заострены | Верхушка почки | Средняя длина | от первичного побега до целого бутона | |||||||||||
| d.d. | д.х. | о.е. h −1 | Основание | Половина | Верх | Основание бутона | Верх бутона | Полностью видно | Частично видно | Не видно | Прим. побег | Целый бутон | ||||||
| В поле | 0,0 | 0,0 | 0,0 | – | – | – | – | – | – 9068 – 9068 | 2.0 | 7,5 | 0,3 | ||||||
| Предварительный день 5 | 0,0 | 0,0 | 168,6 | – | – | – | – | – | – | – | – | 2,0 | 8,0 | 0,3 | ||||
| Предварительный день 7 | 0,0 | 0,0 | 233,2 | + | – | – | – | – | – | – | – | – | – | 3.9 | 12,5 | 0,3 | ||
| Форсирование дней | ||||||||||||||||||
| 1 | 15.9 | 382.6 | 647.7 | + | – | – | – | – 9068 | – | – | 3,9 | 11,4 | 0,3 | |||||
| 2 | 31,8 | 762,6 | 1059,2 | ++ | – | – | – | × | – | – | 2.0 | 8,4 | 0,3 | |||||
| 3 | 47,5 | 1139,2 | 1466,6 | ++ | – | – | – | – | – | – | – | 2,0 | 8,1 | 0,3 | ||||
| 4 | 63,3 | 1518,8 | 1877,4 | + | × | – | – | – | – | – | 90702.6 | 6,6 | 0,3 | |||||
| 5 2,4 | 79,0 | 1895,2 | 2284,4 | – | × | – | × | × | – | 8,7 | 0,3 | |||||||
| 6 | 94,8 | 2274,0 | 2694,4 | – | × | – | – | × | – | × | – 2 .5 | 7,8 | 0,4 | |||||
| 7 | 110,7 | 2656,7 | 3109,2 | × | – | × | – | × | – | × | – | 7,1 | 0,4 | |||||
| 8 | 126,7 | 3039,7 | 3524,4 | × | – | × | × 6 | (8) | 3.4 | 7,5 | 0,5 | |||||||
| 9 | 142,6 | 3421,4 | 3937,6 | × | × | × | × | 2,9 | 8,2 | 0,4 | ||||||||
| 10 | 158,5 | 3803,5 | 4351,6 | × | × | 3.1 | 9,5 | 0,4 | ||||||||||
| 11 | 174,2 | 4180,9 | 4759,9 | × | × | × | 0,5 | |||||||||||
| 12 | 190,1 | 4562,6 | 5173,4 | × | × | .1 | 9,8 | 0,4 | ||||||||||
| 13 | 205,9 | 4940,8 | 5582,7 | × | 0,5 | |||||||||||||
| 14 | 221,8 | 5322,0 | 5995,8 | × | × | 1 | 9,5 | 0,5 | ||||||||||
| 15 | 237,7 | 5704,7 | 6410,6 | × | × | 0,5 | ||||||||||||
| 19 | 297,9 | 7149,8 | 7967.1 | × | × | 7 | 90.310,1 | 0,8 | ||||||||||
| 20 | 312,9 | 7509,5 | 8354,2 | × | × | 0,8 | ||||||||||||
| Количество бутонов на образец | 3 | 3 | 10 | 10 | 2 | 2 | ||||||||||||
| Отбор образцов | Первичные иглы заострены на | Верхушка бутона | Средняя длина (мм) | Среднее отношение первичного побега к целому бутону | ||||||||||||||
| d.d. | д.х. | о.е. h −1 | Основание | Половина | Верх | Основание бутона | Верх бутона | Полностью видно | Частично видно | Не видно | Прим. побег | Целый бутон | ||||||
| В поле | 0,0 | 0,0 | 0,0 | – | – | – | – | – | – 9068 – 9068 | 2.0 | 7,5 | 0,3 | ||||||
| Предварительный день 5 | 0,0 | 0,0 | 168,6 | – | – | – | – | – | – | – | – | 2,0 | 8,0 | 0,3 | ||||
| Предварительный день 7 | 0,0 | 0,0 | 233,2 | + | – | – | – | – | – | – | – | – | – | 3.9 | 12,5 | 0,3 | ||
| Форсирование дней | ||||||||||||||||||
| 1 | 15.9 | 382.6 | 647.7 | + | – | – | – | – 9068 | – | – | 3,9 | 11,4 | 0,3 | |||||
| 2 | 31,8 | 762,6 | 1059,2 | ++ | – | – | – | × | – | – | 2.0 | 8,4 | 0,3 | |||||
| 3 | 47,5 | 1139,2 | 1466,6 | ++ | – | – | – | – | – | – | – | 2,0 | 8,1 | 0,3 | ||||
| 4 | 63,3 | 1518,8 | 1877,4 | + | × | – | – | – | – | – | 90702.6 | 6,6 | 0,3 | |||||
| 5 2,4 | 79,0 | 1895,2 | 2284,4 | – | × | – | × | × | – | 8,7 | 0,3 | |||||||
| 6 | 94,8 | 2274,0 | 2694,4 | – | × | – | – | × | – | × | – 2 .5 | 7,8 | 0,4 | |||||
| 7 | 110,7 | 2656,7 | 3109,2 | × | – | × | – | × | – | × | – | 7,1 | 0,4 | |||||
| 8 | 126,7 | 3039,7 | 3524,4 | × | – | × | × 6 | (8) | 3.4 | 7,5 | 0,5 | |||||||
| 9 | 142,6 | 3421,4 | 3937,6 | × | × | × | × | 2,9 | 8,2 | 0,4 | ||||||||
| 10 | 158,5 | 3803,5 | 4351,6 | × | × | 3.1 | 9,5 | 0,4 | ||||||||||
| 11 | 174,2 | 4180,9 | 4759,9 | × | × | × | 0,5 | |||||||||||
| 12 | 190,1 | 4562,6 | 5173,4 | × | × | .1 | 9,8 | 0,4 | ||||||||||
| 13 | 205,9 | 4940,8 | 5582,7 | × | 0,5 | |||||||||||||
| 14 | 221,8 | 5322,0 | 5995,8 | × | × | 1 | 9,5 | 0,5 | ||||||||||
| 15 | 237,7 | 5704,7 | 6410,6 | × | × | 0,5 | ||||||||||||
| 19 | 297,9 | 7149,8 | 7967.1 | × | × | 7 | 90.310,1 | 0,8 | ||||||||||
| 20 | 312,9 | 7509,5 | 8354,2 | × | × | 0,8 | ||||||||||||
| Количество бутонов на образец | 3 | 3 | 10 | 10 | 2 | 2 | ||||||||||||
Процедура отбора образцов конечных и мутовчатых почек на образце и ветви ануса Морфологические анализы и измерения длины показаны в схематической форме на Рисунке 2.Образцы для анатомического анализа отбирались с 17 января до 6-го дня воздействия, а для всех других анализов – 20 раз с 17 января до 20-го дня воздействия, когда исследование было прекращено. Фиксацию анатомических образцов проводили, как описано Kivimäenpää et al. (2004). Продольные срезы двух конечных зачатков, а также поперечные срезы от верхушки одной зачатка оборота (рис. 2) были разрезаны на ультратоме LKB 2128 (Bromma, Швеция). Срезы толщиной 1,5 мкм окрашивали, как описано Kivimäenpää et al.(2004), а возможные анатомические и клеточные изменения наблюдали под световым микроскопом (Leica DM2500, Германия) при увеличении 100–400 × и фотографировали с помощью цифровой камеры (Leica CD Camera, Швейцария). Adobe Photoshop (версия 6.0) использовался для измерения и анализа цифровых изображений.
Рис. 2.
Схематическая диаграмма верхней части ответвления образца. Терминальные почки (Tb) и почки мутовки (Wb) были взяты для анализа с конечного побега ветви и с четырех ближайших к нему боковых побегов.Линии показывают, как пять кончиков побегов были отрезаны от ветки. Черный цвет двух конечных почек, а также черный цвет одного мутовчатого почек указывает на части почек, взятые для анатомических исследований (химически фиксированные образцы). Белые части этих двух конечных почек использовали для измерения длины роста первичного побега и всей почки. Конечные зачатки, отмеченные звездочками, были недавно разрезаны в продольном направлении для анатомического анализа трахеид. Немаркированные бутоны мутовок были удалены с их чешуек и сфотографированы для морфологического анализа примордиальных игл и вершины.Морфология зачатков хвои всех остальных почек также наблюдалась при подготовке почек для других анализов.
Рис. 2.
Схематическая диаграмма верхней части ветви образца. Терминальные почки (Tb) и почки мутовки (Wb) были взяты для анализа с конечного побега ветви и с четырех ближайших к нему боковых побегов. Линии показывают, как пять кончиков побегов были отрезаны от ветки. Черный цвет двух конечных почек, а также черный цвет одного мутовчатого почек указывает на части почек, взятые для анатомических исследований (химически фиксированные образцы).Белые части этих двух конечных почек использовали для измерения длины роста первичного побега и всей почки. Конечные зачатки, отмеченные звездочками, были недавно разрезаны в продольном направлении для анатомического анализа трахеид. Немаркированные бутоны мутовок были удалены с их чешуек и сфотографированы для морфологического анализа примордиальных игл и вершины. Морфология зачатков хвои всех остальных почек также наблюдалась при подготовке почек для других анализов.
Длину всей почки и длины первичного побега двух терминальных почек (рис. 2) измеряли от самой нижней части пластинки колленхимы до вершины почки или первичного побега под стереомикроскопом (Wild, Швейцария) на 60-кратное увеличение (см. Рис. 3А). Рассчитывали отношение длины первичного побега к длине всей почки. Продольные свежие срезы были вырезаны из оставшихся трех терминальных почек (рис. 2) лезвием бритвы под стереомикроскопом.Свежесрезанные срезы окрашивали 1% толуидиновым синим в течение 2 минут и исследовали под световым микроскопом при увеличении 100 и 400 на предмет развития трахеид и других возможных изменений.
Рис. 3.
(A) Продольный разрез вегетативной почки ели, отобранный в поле 17 января 2007 г. Первичный побег составляет около одной трети общей длины почки. Линия на бутоне показывает точку измерения первичного побега и всей почки.Чешуйки почек (Bs) компактные и блестящие на внешней поверхности, но нежные и белые вокруг первичного побега. Пруток = 1 мм. (B) Верхняя поверхность первичного побега из A. Первичные иглы плотно прижаты к первичному побегу, и их кончики все тупые (C), что придает кончикам округлый вид, если смотреть вокруг голой звездообразной вершины наверху. бутон. Прутки в B = 0,5 мм и в C = 0,2 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 3.
(A) Продольный разрез вегетативной почки ели, отобранный в поле 17 января 2007 г. Первичный побег составляет около одной трети общей длины почки. Линия на бутоне показывает точку измерения первичного побега и всей почки. Чешуйки почек (Bs) компактные и блестящие на внешней поверхности, но нежные и белые вокруг первичного побега. Пруток = 1 мм. (B) Верхняя поверхность первичного побега из A. Первичные иглы плотно прижаты к первичному побегу, и их кончики все тупые (C), что придает кончикам округлый вид, если смотреть вокруг голой звездообразной вершины наверху. бутон.Прутки в B = 0,5 мм и в C = 0,2 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Из четырех боковых побегов по одной зачатке мутовки на побег (рис. 2) был удален из их чешуек почек, и верхняя поверхность зеленых первичных зачатков наблюдалась под стереомикроскопом при 60–120-кратном увеличении под разными углами, чтобы определить фазы развития зачатков хвои и появление верхушки. Первоначальные иглы и верхушка также наблюдались на всех других почек, когда они готовились к другим исследованиям.
Результаты
Суммарное охлаждение с 1 сентября по 17 января, которое было днем отбора проб на полях, составило 1237 единиц охлаждения (ch.u. h −1 ) (Sarvas 1974) (Рисунок 1). Почки, отобранные в поле, имели типичную структуру покоя. Среднее отношение длины первичного побега к общей длине почки составляло 0,3 (рис. 3А; таблица 1). Чешуйки почек были компактными, а внешняя поверхность блестела, тогда как внутренние чешуйки рядом с первичным побегом были нежными и живыми (рис. 3А).Первичные иглы, которые были плотно прижаты к первичному побегу и окружали голую звездообразную вершину на верхушке почки, были тупыми, придавая кончикам округлый вид (рис. 3B и C). Под световым микроскопом меристематические клетки, которые характеризуются большим ядром и гладкой цитоплазмой без вакуоли, доминировали везде, за исключением прокамбиальных рядов рядом с самыми нижними зачатками игл и некоторых дубильных вакуолизированных клеток в примордиальном побеге (рис. 4А). .Центральные материнские клетки в верхушке имели маленькие липидные капельки, которых не было в других клетках верхушки.
Рисунок 4.
(A) Продольный разрез химически закрепленной вегетативной почки, взятой на 2-й день выгонки. Прокамбиальные ряды, характеризующиеся удлиненными клетками (звездочки), можно увидеть на уровне самых нижних примордиальных игл (N). Накопление липидных капель (отмеченная область на A увеличена на B) можно увидеть в меристематических клетках примордиальных игл и в меристеме между прокамбиальным рядом и примордиальными иглами.Обратите внимание на дубильные (Т) клетки в первичном побеге. Штанги в A = 50 мкм и в B = 10 мкм. (C) Продольный фиксированный срез от основания вегетативной почки, отобранный на 6-й день выгонки. Накопление липидов больше не различимо, но дифференциация первичной проводящей ткани (Ct) и игл (N) увеличилась по сравнению со 2-м днем. = 50 мкм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 4.
(A) Продольный разрез химически закрепленной вегетативной почки, взятой на 2-й день выгонки.Прокамбиальные ряды, характеризующиеся удлиненными клетками (звездочками), видны на уровне самых нижних примордиальных игл (N). Накопление липидных капель (отмеченная область на A увеличена на B) можно увидеть в меристематических клетках примордиальных игл и в меристеме между прокамбиальным рядом и примордиальными иглами. Обратите внимание на дубильные (Т) клетки в первичном побеге. Штанги в A = 50 мкм и в B = 10 мкм. (C) Продольный фиксированный срез от основания вегетативной почки, отобранный на 6-й день выгонки.Накопление липидов больше не различимо, но дифференциация первичной проводящей ткани (Ct) и игл (N) увеличилась по сравнению со вторым днем. Полоса = 50 мкм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
На уровне светового микроскопа первым наблюдаемым изменением было увеличение количества липидов в фиксированных образцах, собранных на 7-й день перед форсированием (Таблица 1). В это время сумма градус-дней (д.д.) и сумма градус-часов (д.ч.) оставались равными нулю, тогда как сумма за период составляла 233 о.е. h −1 . Увеличенное количество и размер липидных капель впервые были замечены в меристеме между прокамбиальным рядом и нижними примордиальными иглами (рис. 4A и B). В максимуме во второй и третий дни выгонки (таблица 1) липидные капли можно было увидеть почти повсюду в меристематических клетках вплоть до области верхушки почек. На 5-й и 6-й дни принуждения явление больше не было видимым, но дифференциация первичной проводящей ткани и тканей иглы стала четкой (рис. 4С).Первые маленькие трахеиды в первичной проводящей ткани были обнаружены рядом с самыми нижними примордиальными иглами в свежесрезанных продольных образцах на 4-й, 5-й и 6-й дни воздействия (Рисунок 5A и B; Таблица 1). На 7-й и 8-й дни трахеиды были длинными, хорошо развитыми и вытянутыми до половины длины первичного побега, а с 9-го дня и далее до примордиальных игл рядом с верхушкой (Рисунок 6A; Таблица 1).
Рис. 5.
(A и B) Свежесрезанные продольные срезы первичного побега вегетативной почки ели, взятые на 5-й день выгонки, когда можно было различить первые трахеиды (звездочки на B) (отмеченная область на A увеличена в B).Прутки в A = 0,2 мм и в B = 0,020 мм. (C) Верхняя поверхность первичного побега из вегетативной почки ели, отобранная на 5-й день выгонки, показывает, как первые первичные иглы у основания почки стали заостренными (стрелки). Пруток = 0,5 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 5.
(A и B) Свежесрезанные продольные срезы первичного побега вегетативной почки ели, отобранные на 5-й день выгонки, когда можно было различить первые трахеиды (звездочки на B) (отмеченная область на A обозначена увеличено в B).Прутки в A = 0,2 мм и в B = 0,020 мм. (C) Верхняя поверхность первичного побега из вегетативной почки ели, отобранная на 5-й день выгонки, показывает, как первые первичные иглы у основания почки стали заостренными (стрелки). Пруток = 0,5 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 6.
(A) Свежесрезанный продольный разрез первичного побега вегетативной почки ели, отобранный на 10-й день выгонки.Хорошо развитая проводящая ткань (стрелки) видна до верхушки почки. Пруток = 0,5 мм. (B) Верхняя часть первичного побега из почек, отобранная на 12-й день выгонки, когда самые верхние первичные иглы стали заостренными и покрыли вершину. Пруток = 0,5 мм. (C) Продольный разрез вегетативной почки ели, отобранный на 13-й день выгонки. Первичный побег удлинен, а чешуйки почек, если смотреть с внешней поверхности на D, слегка раздвинуты. Прутки в C и D = 1 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 6.
(A) Свежесрезанный продольный разрез первичного побега вегетативной почки ели, отобранный в 10-й день выгонки. Хорошо развитая проводящая ткань (стрелки) видна до верхушки почки. Пруток = 0,5 мм. (B) Верхняя часть первичного побега из почек, отобранная на 12-й день выгонки, когда самые верхние первичные иглы стали заостренными и покрыли вершину. Пруток = 0,5 мм. (C) Продольный разрез вегетативной почки ели, отобранный на 13-й день выгонки. Первичный побег удлинен, а чешуйки почек, если смотреть с внешней поверхности на D, слегка раздвинуты.Прутки в C и D = 1 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
На стереомикроскопическом уровне первые изменения почек были замечены на 5-й день выгонки, когда кончики самых нижних примордиальных игл стали заостренными (Рисунок 5C; Таблица 1). До этого момента практически не происходило удлинения зачатков побегов (таблица 1), а чешуйки почек оставались компактными и блестящими, как зимой (см. Рис. 3А). Начиная с 8-го дня, примордиальные иглы почти во всех почек начали заостряться даже на вершине первичного побега, и, следовательно, вершина начала частично покрываться заостренными первичными иглами (Таблица 1).До 11-го дня чешуя почек была еще более или менее в зимнем состоянии. Из-за того, что все первичные иголки выросли в длину, верхушка больше не была видна в большинстве почек к 12-му дню и ни в одной из почек с 13-го дня и далее (Рисунок 6B; Таблица 1). На 12-й день чешуйки почек начали ослабевать и слегка раздвинулись на 13-й день, когда первичные побеги также составляли около половины общей длины вегетативной почки (рис. 6C и D; таблица 1). В 19 и 20 дни (313 д.д., 7510 д.п., 8354 р.u. h -1 ; Таблица 1), примордиальные побеги составляли 80% или более от общей длины почки, а чешуйки почки сильно торчали (Рис. 7A и B) и были очень рыхлыми (Рис. 7C), или распад почки завершился (Рис. 7D). . Маленькие чешуйки вновь появившейся почки можно было увидеть на вершине развивающейся «старой» почки под удлиняющимися первичными иглами с 12 дня (см. Рис. 7A).
Рис. 7.
(A и B) Образцы вегетативных почек ели, взятые на 19-й день выгонки.(A) Первичный побег почти такой же длины, как и вся почка, а наверху видна недавно появившаяся почка (звездочка). (B) Чешуйки почек сильно выступают. (C) Новые иголки (звездочки) можно различить по распущенным чешуйкам почек и (D) без чешуек после распускания почек. Штанги на всех рисунках = 1 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Рис. 7.
(A и B) Вегетативная почка ели, отобранная на 19-й день выгонки. (A) Первоначальный побег почти такой же длины, как и вся почка, и недавно появившаяся почка (звездочка) видна вверху.(B) Чешуйки почек сильно выступают. (C) Новые иголки (звездочки) можно различить по распущенным чешуйкам почек и (D) без чешуек после распускания почек. Штанги на всех рисунках = 1 мм. Эта цифра отображается в цвете в онлайн-версии Tree Physiology .
Обсуждение
Структура спящих вегетативных почек (например, небольшой размер первичного побега по сравнению с размером всей почки и меристематическая природа почти всех клеток почек), наблюдаемая в этом исследовании 17 января в полевых условиях, была аналогична описанной. в других местах почек ели обыкновенной отбирали только с веток (Hejnowicz and Obarska 1995, Bílková et al.1999, Guzicka and Woźny 2003) или с ветвей и верхних побегов молодых и взрослых деревьев (Sutinen et al. 2006, 2007). С 1 сентября по 17 января величина накопленного переохлаждения составила 1237 ч.е. h −1 . Поскольку накопленное охлаждение, необходимое для высвобождения эндодормальности и начала онтогенетического развития в направлении распускания почек, составляет 420 ch.u. h −1 для ели европейской (Sarvas 1974), очевидно, что почки на ветвях, собранных для наших исследований выгонки, уже находились в стадии экопадения, т.е.е., готовые расти в благоприятных условиях.
Крахмал, углеводы и жиры – важнейшие запасы энергии растений. Хвоя и внутренняя кора ветвей (Судачкова и др., 2004), а также почки (Липавска и др., 2000) содержат такие запасы энергии. Таким образом, бутоны самодостаточны в отношении питания, по крайней мере, на ранних стадиях развития. То же самое было и с нашими отделившимися ветвями, потому что можно было проследить за развитием почек до тех пор, пока они не лопнут. С поздней осени до состояния позднего покоя концентрации сахаров (Lipavská et al.2000) и липиды (Jordy 2004) в почках находятся на самом высоком уровне. По мере приближения к распусканию почек концентрация сахаров и липидов в почках / хвое уменьшается, что указывает на использование этих запасных веществ для развития почек (Jordy 2004, Sudachkova et al. 2004). В этом исследовании временное увеличение размера и количества липидных капель в течение первых четырех дней нагнетания, по-видимому, было связано с обеспечением энергией для развития первых трахеидов, которые появились на 4-й день нагнетания.Накопление крахмала в этом исследовании не было обнаружено, что может быть связано с отсутствием крахмальных зерен или крошечным размером крахмальных зерен, которые нельзя было различить на световом микроскопическом уровне в фиксированном материале в течение первых шести дней нагнетания. Необходимы дальнейшие исследования в этой области, чтобы получить более полное представление о годовой динамике запасов соединений и их появлении в спящих и развивающихся почек ели в бореальной зоне.
Увеличение липидов, наблюдаемое только на клеточном уровне в химически закрепленных почках, произошло, когда сумма единиц периода достигла 233 p.u. h −1 , но сумма температур ( T ≥ +5 ° C) все еще была равна нулю. Таким образом, согласно этому исследованию, накопление единиц периода было бы лучшим индикатором ранних стадий развития почек, чем обычная сумма температур. Однако из-за отсутствия количественных знаний о динамике липидов в почках в полевых условиях истинная взаимосвязь между накоплением липидов и температурой в развитии бутонов остается только на гипотетическом уровне.
Развитие трахеид было тесно синхронизировано с морфологическим изменением от закругленных к заостренным кончикам примордиальных игл.К 11-му дню выгонки заостренные иголки на всех почках до вершины были еще частично видны. Первые внешние видимые изменения были замечены на 12-й день выгонки, когда начали торчать чешуйки почек. Подобная ранняя фаза развития почек (стадия B1) у P. abies была описана ранее Slaney et al. (2007). Морфологическое изменение хвои завершилось на 13-й день выгонки, т.е. на пятый день после фазы, когда трахеиды достигли вершины развивающейся почки.К этому времени удлиненная хвоя покрывала верхушку, а зачаток вырос до половины всей длины почки. О важности трахеид для распускания почек и быстрого удлинения сообщалось ранее у P. abies (de Faÿ et al. 2000). С 12-го по 20-й день выгонки расщепление и опадание чешуи почек были аналогичны тем, которые описаны ранее для ели европейской Slaney et al. (2007).
Сумма температур для наблюдаемого распада почек (распускание чешуек почек и появление кончиков игл) ели европейской, происходящей из северных широт (58 ° N – 68 ° N), варьируется от 129 до 189 дней.d., в зависимости, например, от широты и высоты происхождения семян (Beuker 1994, Hannerz 1999, Slaney et al. 2007). В форсированных условиях нашего исследования сумма температур 190 д.д. была достигнута на 12-й день, когда все иголки стали заостренными, стало очевидным удлинение первичного побега и начали расшатываться чешуйки почек. Ближе к распусканию бутонов (20-е сутки) сумма температур составила 313 сут. Распускание почек у ели европейской в исследовании Slaney et al. (2007) заняло 40–50 дней (по оценке, приведенной на Рисунке 5 в Slaney et al.2007), тогда как в условиях выгонки в этом исследовании период между распусканием почек и первыми видимыми изменениями в чешуях почек длился 9 дней. Из вышеизложенного ясно, что сумма температур, накопленная за период выгонки, завышает требования к сумме температур для биологических фаз развития почек.
В нашем эксперименте с одним деревом различия между бутонами наблюдались только во времени фаз, когда первичные иглы заострялись на верхушке почек и когда верхушка была полностью покрыта удлиненными первичными иглами.Необходим более полный полевой материал для определения вариаций в скорости развития почек внутри и между деревьями, а также между деревьями разного возраста. Также необходимы дальнейшие исследования в этой области, чтобы определить, как наблюдаемые здесь фазы развиваются при температурах в полевых условиях, и какие из наблюдаемых фаз могут быть использованы на практике в обширных исследованиях, а также для оценки и улучшения моделей. Хотя сроки морфологических изменений в пределах одного дерева, между отдельными деревьями и между деревьями с разным семенным происхождением различаются, структура и последовательность фаз развития почек у ели европейской, скорее всего, следует той же схеме, что и здесь.
В заключение, первым признаком развития почек было увеличение размера и количества липидных капель при очень низком суммарном значении температуры. Затем последовало развитие трахеид от основания до верхушки почек, что было тесно синхронизировано с морфологическим изменением зачатых игл с округлых на заостренные. Основываясь на морфологических изменениях хвои и удлинении первичного побега, развитие почек можно разделить на следующие четыре фазы: (i) заостренные заостренные иглы в основании почки, (ii) самые верхние первичные иглы. заострены и частично покрывают ранее обнаженную верхушку, (iii) все иглы начали удлиняться и, таким образом, полностью покрывают верхушку, и (iv) видимое удлинение бутона.Морфологические изменения примордиальных игл на остроконечные (фазы i и ii) произошли без каких-либо внешне видимых изменений почек. Фаза iii соответствует началу видимого снаружи развития почек, а фаза iv соответствует видимому удлинению почек. Таким образом, фазы i и ii потенциально могут быть использованы в будущих полевых исследованиях для тестирования и улучшения фенологических моделей вместе с наблюдениями за видимым распусканием почек. Однако суммарные значения температуры, необходимые для фаз развития почек, в поле, скорее всего, будут ниже, чем в условиях выгонки, использованных в этом исследовании.
Мы благодарим Сейю Репо, Яану Риссанен, Сакари Сильвеннойнен и Йоуко Лехто за их помощь в полевых условиях и в лаборатории. Это исследование финансировалось Финским культурным фондом и Фондом Эллы и Георга Эрнроот.
Список литературы
,,,,.Изменения европейской весенней фенологии
,Int. J. Climatol.
,2002
, т.22
(стр.1727
–1738
).Адаптация к климатическим изменениям сроков распускания почек в популяциях Pinus sylvestris L.и Picea abies (L.) Karst.
,Tree Physiol.
,1994
, т.14
(стр.961
–970
),,.Гистохимическое определение и анализ изображений неспецифической активности эстеразы и количества полифенолов во время развития однолетних почек у ели европейской
,J. Exp. Бот.
,1999
, т.35
(стр.1129
–1138
),,,.Парниковый эффект, изменение климата и экосистемы
,СФЕРА ПРИМЕНЕНИЯ 29
,1986
Чичестер, Великобритания
John Wiley and Sons
541
.Типы лесов и их значение
,Acta For. Фенн.
,1949
, т.56
(стр.1
–71
),.Фенология распускания почек у пихты Дугласа, связанная с происхождением, фотопериодом, охлаждением и температурой промывки
,Бот. Газ.
,1975
, т.136
(стр.290
–298
),.Термическое время, холодные дни и прогноз распускания почек у Picea sitchensis
,J.Прил. Ecol.
,1983
, т.23
(стр.177
–191
),,. ,SILMUSCEN и CLIGEN руководство пользователя; руководящие принципы построения климатических сценариев и использования стохастического погодного генератора в Финской программе исследований изменения климата (SILMU)
,1995
Публикации Академии Финляндии 5/95
62
.Обзор литературы по покою почек древесных растений
,Мед.ван де Ландбоухогеш. Вагенинген, Нидерланды
,1953
, т.53
(стр.1
–23
),,.Связанные с водой явления в озимых почках и ветках Picea abies L. (Karst.) До распускания почек: биологическое, гистологическое и ЯМР исследование
,Ann. Бот.
,2000
, т.86
(стр.1097
–1107
),,,,. ,.Модель ступени роста (° GS) и холодовая акклиматизация древесных растений умеренного пояса
,Холодостойкость растений и морозостойкость.Механизмы и последствия для сельскохозяйственных культур
,1982
, vol.Т. 2
Нью-Йорк
Academic Press
(стр.93
–116
),.Изменения ультраструктуры в середине зимы в вегетативных зародышевых побегах ели европейской [ Picea abies (L.) Karst.]
,Ann. Для. Sci.
,2003
, т.60
(стр.461
–466
),,,.Гомолог Т локуса цветения ели европейской участвует в контроле ритма роста у хвойных деревьев
,Plant Physiol.
,2007
, т.144
(стр.248
–257
),,.Влияние покоя и факторов окружающей среды на время распускания почек у Betula pendula
,Tree Physiol.
,1998
, т.18
(стр.707
–712
).Оценка температурных моделей для прогнозирования распускания почек у ели обыкновенной
,Кан. J. For. Res.
,1999
, т.29
(стр.9
–19
).Моделирование высвобождения покоя почек у деревьев из регионов с прохладным и умеренным климатом
,Acta For. Фенн.
,1990
, т.213
(стр.1
–47
).Повышает ли потепление климата риск повреждения северных деревьев морозом?
,Среда растительных клеток.
,1991
, т.14
(стр.449
–454
).Влияние климатических изменений на деревья из регионов с прохладным и умеренным климатом: экофизиологический подход к моделированию фенологии распускания почек
,Can.J. Bot.
,1995
, т.73
(стр.183
–199
). ,,.Физиологические аспекты климатической адаптации растений с особым упором на среду высоких широт
,Растениеводство на Севере
,1985
Тромсе
Norwegian University Press
(стр.1
–22
).Продолжительность светового дня и термическое время реакции распускания почек во время выхода из покоя у некоторых северных лиственных деревьев
,Physiol.Растение.
,1993
, т.88
(стр.531
–540
).Для восстановления покоя почек бука ( Fagus sylvatica ) требуется как охлаждение, так и долгие дни
,Physiol. Растение.
,1993
, т.89
(стр.187
–191
).Высокая осенняя температура задерживает весеннее распускание почек на северных деревьях, уравновешивая эффект потепления климата
,Tree Physiol.
,2003
, т.23
(стр.931
–936
),.Строение и развитие вегетативных почек от нижней кроны Picea abies
,Ann. Sci. Для.
,1995
, т.52
(стр.433
–447
),.Ограничения роста бореальных лесов
,Природа
,2000
, т.405
(стр.904
–905
).Сезонная изменчивость органогенетической активности и распределения резервов в верхушке побега Pinus pinaster Ait.
,Ann. Бот.
,2004
, т.93
(стр.25
–37
),,.Деакклимация и реакклимация морозостойких растений: текущее понимание и новые концепции
,Plant Sci.
,2006
, т.171
(стр.3
–16
),,.Расчеты морозостойкости сосны обыкновенной при потеплении климата в бореальных условиях
,Экол. Прил.
,1995
, т.5
(стр.42
–52
),,,,.Использование световой и электронной микроскопии для оценки воздействия озона на хвою ели европейской
,Environ. Загрязнение.
,2004
, т.127
(стр.441
–453
),.Моделирование температурных эффектов при разрыве покоя у кизила краснолистного ( Cornus sericea L.)
,Ann. Бот.
,1983
, т.52
(стр.205
–215
),,.Сезонные изменения свойств рибосомных ансамблей вегетативных почек сосны обыкновенной
,Physiol. Растение.
,1987
, т.71
(стр.55
–60
),,,.Эндопара- и экология: физиологическая терминология и классификация для исследования покоя
,Hortic. Sci.
,1987
, т.22
(стр.371
–377
),,,.Прогнозирование весенней фенологии и риска повреждения морозами Betula sp.при климатическом потеплении: сравнение двух моделей
,Tree Physiol.
,2000
, т.20
(стр.1175
–1182
),,.Годовая динамика содержания неструктурных сахаридов в контексте структурного развития вегетативных почек ели европейской
,J. Plant Physiol.
,2000
, т.157
(стр.365
–373
),,.Влияние фотопериода и температуры на время распускания почек у ели обыкновенной ( Picea abies )
,Tree Physiol.
,1998
, т.18
(стр.811
–816
),,.Влияние накопленной продолжительности светового периода на распускание почек у ели обыкновенной ( Picea abies ) разного возраста
,Silva Fenn.
,2001
, т.35
(стр.111
–117
).Покой деревьев зимой
,Наука
,1971
, т.171
(стр.29
–36
).Меристемы, рост и развитие древесных растений
,USDA For. Серв. Tech. Бык.
,1963
, т.1293
214
.Покой древесных растений
,Ann. Rev. Plant Physiol.
,1954
, т.5
(стр.183
–204
).Исследования годового цикла развития деревьев леса. Активный период
,Комм. Inst. Для. Фенн.
,1972
, т.76
(стр.1
–110
).Исследования годового цикла развития лесных деревьев II. Осенний и зимний покой
,Comm. Inst. Для. Фенн.
,1974
, т.84
(стр.1
–101
).Парниковый эффект: наука и политика
,Science
,1989
, vol.243
(стр.771
–781
),,,.Влияние повышенной концентрации углекислого газа и температуры на распускание почек и рост побегов северной ели европейской
,Tree Physiol.
,2007
, т.27
(стр.301
–312
),.Химическая регуляция фаз покоя развития почек
,Am. J. Bot.
,1964
, т.51
(стр.1002
–1012
).Ультраструктура мезофилла сосны обыкновенной и ели европейской: сезонные колебания и молярность фиксирующего буфера
,Protoplasma
,1980
, vol.103
(стр.241
–252
),,,.Годовая динамика активности резервных соединений и гидролитических ферментов в тканях Pinus sylvestris L. и Larix sibirica Ledeb. Метаболизм резервных соединений в тканях хвойных пород сибирских
, евразийскихJ. For. Res.
,2004
, т.7–1
(стр.1
–10
),,.Нарушения развития почек и хвои зрелой ели европейской, Picea abies (L.) Karst., В зависимости от содержания бора
,Деревья
,2006
, vol.20
(стр.559
–570
),,.Влияет ли время внесения бора на структуру хвои и почек у сеянцев сосны обыкновенной и ели европейской?
,Деревья
,2007
, т.21
(стр.661
–670
),,.Неструктурный углеводный статус почек ели европейской в контексте структурного развития однолетних почек под воздействием кислотного загрязнения
,Environ. Exp. Бот.
,2000
, т.43
(стр.253
–265
).Холодостойкость и травмы древесных растений
,Science
,1970
, vol.169
(стр.1269
–1278
),,,,,.Экспрессия дегидринов, связанная со временем распускания почек у ели европейской
,Planta
,2008
, vol.228
(стр.459
–472
)© Автор 2009. Опубликовано Oxford University Press.Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]
.границ | Изменения метаболитов в почек и хвои хвойных во время принудительного распускания почек у ели обыкновенной (Picea abies) и пихты обыкновенной (Abies alba)
Введение
За последние 150 лет сжигание богатого углеродом нефтяного топлива и индустриализация привели к увеличению концентрации CO 2 примерно на 43% с доиндустриальной концентрации около 280 ppm до почти 400 ppm сегодня (IPCC, 2007).Это привело к хорошо задокументированному повышению глобальной температуры и связанных с ней климатических моделей, причем потепление, как правило, сильнее в более высоких широтах. Ожидается, что эти тенденции сохранятся. Одним из возможных последствий глобального потепления является нарушение фенологии завязывания почек, покоя и распускания почек у деревьев и других растений в лесных экосистемах с умеренно холодным климатом и бореальных лесах.
В глобальном масштабе леса представляют собой крупнейший наземный пул углерода на Земле, большая часть которого хранится в северных бореальных древесных лесах (Dixon et al., 1994). Хвойные и смешанные леса занимают площадь около 2,5 × 10 6 км 2 и действуют как поглотители углерода (Breymeyer et al., 1996). Изучение последствий потепления климата на некоторых бореальных хвойных деревьях выявило изменения в углеродном балансе между фотосинтезом и дыханием (Kirschbaum, 2000). Такие изменения могут повлиять на способ развития растений (Norby and Luo, 2004) и могут отрицательно повлиять на метаболические процессы в деревьях (Lavender and Silim, 1987), особенно в периоды, когда деревья подвергаются критическим физиологическим процессам, таким как образование почек или распускание почек. .
Время распускания почек имеет фундаментальное значение для выживания и роста деревьев в умеренных и северных регионах мира (Sakai and Larcher, 1987). Преждевременное распускание почек зимой или весной может подвергнуть развивающуюся ткань стебля и листьев воздействию мороза, в то время как отсроченное распускание почек может сократить вегетационный период. В бутонах цветов или шишек эти изменения могут повлиять на время и успех опыления, а также на время вегетационного периода, доступное для развития плодов, шишек и семян.
Тремя наиболее важными факторами, контролирующими фенологию покоя у деревьев умеренного и северного климата, являются продолжительность светового дня или фотопериод для индукции покоя (Wareing, 1953; Hannerz, 1999), охлаждающая потребность для перехода от эндодормальности к экодормальности (Vegis, 1964), и потребность в сумме тепла для начала распускания почек.Как правило, повышение температуры вызывает раскрытие почек и ополаскивание дерева (Cannell and Smith, 1984), а сумма тепла, необходимая для раскрытия почек, выраженная как накопленные градусо-дни выше пороговой температуры, уменьшается с увеличением продолжительности охлаждения (Murray et al. al., 1989) до тех пор, пока не будет выполнено требование охлаждения.
Преждевременное распускание почек и цветение в мягкую погоду зимой часто наблюдается у декоративных видов, высаженных за пределами их естественного ареала, а также может наблюдаться в естественных популяциях в результате глобального потепления.Мягкая погода зимой также может привести к потере морозостойкости, увеличивая риск повреждения стеблей, листьев или игл вечнозеленых растений от мороза (Strimbeck et al., 1995). Глобальное потепление, вероятно, увеличит частоту и суровость умеренных погодных явлений в середине зимы, особенно в более высоких широтах, с последующим риском морозного повреждения деревьев и других растений даже в пределах их нынешних естественных ареалов.
Еще одно возможное последствие глобального потепления состоит в том, что потребность в охлаждении может быть не полностью удовлетворена, что приведет к задержке распускания почек (Heide, 1993; Morin et al., 2009). Результаты модели распускания почек у 22 видов лиственных деревьев умеренного пояса, откалиброванные с использованием обширных данных о требованиях к охлаждению и сумме тепла, показывают, что распускание почек у большинства видов может быть увеличено с нескольких дней до более чем 3 недель при умеренном потеплении A2. (+ 1 ° С). В более экстремальном сценарии B2 (+ 3,2 ° C) раскрытие почек может быть отложено во всех или некоторых частях текущего ареала некоторых видов. Более поздние исследования показывают, что возможно или даже вероятно еще более сильное потепление.Таким образом, задержка распускания почек и последующие проблемы развития – реальная возможность для многих северных и умеренных деревьев и других видов растений.
Кроме того, повышенный уровень CO в атмосфере 2 может косвенно влиять на фенологию почек, изменяя их биохимию и физиологию. Например, изменение содержания крахмала или фитогормонов может изменять период покоя и роста, изменяя фенологические закономерности, такие как более раннее цветение или увеличение продолжительности вегетационного периода (Cannell, 1990).У ели европейской ( Picea abies ) время распускания почек было связано с высокой скоростью чистого фотосинтеза, за которым следовало уменьшение количества сахара и повышение содержания крахмальных соединений (Egger et al., 1996). Кроме того, ранней весной перед распусканием почек большое количество углерода выводится из хвои, тогда как после распускания почек концентрация крахмала в тканях уменьшается, а количество неструктурных углеводов и гемицеллюлоз увеличивается вдвое.
Сахар и другие совместимые растворенные вещества обладают криозащитными свойствами и накапливаются в клетках хвойных деревьев во время заморозки.Хотя сахароза является наиболее распространенным растительным сахаром, олигосахариды, такие как раффиноза и стахиоза, тесно связаны с морозостойкостью хвойных деревьев (например, Strimbeck et al., 2007). Основные изменения в метаболизме сахаров происходят в хвое хвои при акклиматизации и дезакклимации (Ларчер, 2003; Ангелчева и др., 2014). Акклимация к холоду обычно тормозит рост, однако повышенный уровень CO 2 вызывает задержку акклиматизации наряду с повышенным риском повреждения растений морозом (Loveys et al., 2006).
У вечнозеленых хвойных растений углеводы и фотосинтаты из прошлогодней хвои поддерживают рост побегов до тех пор, пока не разовьются новые иглы (Hansen and Beck, 1994). Во время распускания почек снабжение углеродом обеспечивается исключительно метаболической активностью накопленных запасов, но считается менее важным для развития новых побегов (Hansen and Beck, 1990). Старая хвоя у Picea abies накапливает большие ресурсы крахмала до распускания почек, которые полностью расходуются во время развития побегов (Senser et al., 1975).
Kalberer et al. (2006) конкретно указывают на необходимость лучшего понимания эволюционной истории и экологического значения фазы деакклимации, обращаясь к передаче сигналов, экспрессии генов и механизмам адаптации, включая метаболизм белков и малых молекул (метаболитов). Платформы и технологии метаболомики с высокой пропускной способностью были внедрены в науку о растениях в последнее десятилетие. В последние годы профилирование метаболитов на основе газовой хроматографии / масс-спектрометрии (ГХ / МС) успешно применялось для анализа молекулярных механизмов, регуляции первичного метаболизма и метаболических сетей у различных родов растений (Fiehn et al., 2000; Ресснер и др., 2000; Lisec et al., 2006), как в отношении акклимации к холоду (например, Cook et al., 2004; Rohloff et al., 2012; Angelcheva et al., 2014), так и фазы дезакклимации (например, Oono et al., 2006; Бюн и др., 2014). Ранние отчеты о метаболомике хвойных пород были сосредоточены на методологии и создании хемометрических алгоритмов для отбора и фенотипирования растений (Robinson et al., 2007; Zou and Tolstikov, 2008), ассоциативной генетике (Eckert et al., 2012) и регенерации растений. соматические культуры (Robinson et al., 2009). В более поздних исследованиях вопросы, связанные с акклиматизацией к холоду у Picea sitchensis (Dauwe et al., 2012) и Picea obovata (Shiryaeva et al., 2012; Angelcheva et al., 2014), были решены с помощью ГХ / МС. профилирование метаболитов на основе. Лишь несколько отчетов приблизились к фазе дезакклимации в отношении повышенных температур, уровней озона (Riikonen et al., 2012) и влияния продолжительности дня на развитие почек и распускание почек (Lee et al., 2014). Всесторонних исследований по регулированию метаболизма во время распускания почек и отвердения у хвойных пород до сих пор нет.
Мы исследовали изменения метаболитов в почках и хвое двух вечнозеленых хвойных деревьев во время искусственной выгонки, чтобы измерить возможные последствия из-за раннего прихода весны. Picea abies L. (Ель обыкновенная) – это бореальный вид, адаптированный к низким температурам, который произрастает по всей Северной Европе, включая Скандинавию и запад России. Abies alba L. (пихта обыкновенная) – вид с умеренным климатом, встречающийся в природе в Центральной Европе. Picea abies является важным промысловым видом на большей части своего ареала, вырастая до 50 м в высоту, а Abies alba представляет собой одно из крупнейших и наиболее важных лесных деревьев в Центральной Европе (Dobrowolska, 1998).Мы использовали профилирование метаболитов на основе ГХ / МС для изучения изменений в пулах преимущественно гидрофильных метаболитов в ткани почек и хвои обоих видов во время распускания почек и отвердения. Мы сообщаем об изменениях в уровнях углеводов, аминокислот, соединений, связанных с циклом Кребса, и вторичных соединений, которые более или менее согласованы между двумя видами, тканями хвои и почек, или и тем, и другим. Наши результаты позволяют по-новому взглянуть на потенциальные эффекты прогнозируемого глобального потепления на фенологические характеристики и биохимические реакции у хвойных деревьев во время деакклимации с учетом различий между видами и типом ткани, а также времени регуляции метаболизма во время распускания почек.
Материалы и методы
Расположение
Образцы растений были собраны в Ботаническом саду Рингве, NTNU, расположенном на полуострове Ладе в 3 милях к востоку от центра Тронхейма, Норвегия (63 ° 25 ‘, 10 ° 25’) и на высоте нескольких метров над уровнем моря. Все образцы деревьев были посажены сразу после создания дендрария в 1973 г. (Strimbeck et al., 2007). Как и в большинстве прибрежных регионов Норвегии, условия были оптимальными для роста растений, исходя из умеренности климата из-за Североатлантического течения, ответвления потока, протекающего в сторону полюсов.Согласно норвежской климатической системе, местный климат классифицируется как южно-бореальный и умеренно океанический (Moen, 1999) и, согласно более обобщенному способу классификации глобального климата, описывается как умеренный океанический климат.
Дизайн отбора проб и инкубация
Ветки с бутонами видов Picea abies и Abies alba были собраны в середине марта 2010 г. (всего 216 образцов), по 4 дерева на вид и 3 образца на дерево.Чтобы оценивать симптомы распада почек на еженедельной основе, в тепличных условиях закладывали отдельные побеги с иголками и терминальными почками. Всходы помещали в блоки из насыщенной цветочной пены (24 побега на блок) в лотки с прозрачными пластиковыми крышками под кварцевыми галогенными лампами, обеспечивая 200 мкмоль м -2 с -1 PAR. Образцы хранились при постоянной температуре 22 ° C при цикле день / ночь 16/8 часов в течение до 9 недель. Один раз в неделю все оставшиеся образцы оценивали на предмет распускания почек и отбирали одну реплику блока цветочной формы.Образцы бутонов и игл замораживали в жидкости N 2 и хранили при -80 ° C для последующего анализа. Для отбора проб использовали только терминальные почки (2, до 3 бутонов). Образцы почек и хвои (побеги с 1 по 3) с трех одиночных деревьев (деревья с 1 по 3) были объединены ( n = 3), что дало в общей сложности 108 образцов (2 вида × 2 ткани × 3 повторяющихся дерева × 9 недель) для экстракции метаболитов и последующего анализа ГХ / МС.
Подсчет очков Bud Break
В семействе Pinaceae верхние побеги образуются из заранее сформированной почки.Набухание и распускание почек регистрировали один раз в неделю для двух самых верхних почек в соответствии со шкалой от 0 до 8, в которой стадия 0 относилась к спящим, а стадия 8 – к полностью дифференцированным побегам с полноценными базальными иглами (Krutzsch, 1973). ): 0 – спящие почки; 1 , Почки слегка вздутые, хвоя под бутонами загнута назад и наружу; 2 , Почки опухшие, от зеленого до серо-зеленого цвета, чешуя почек еще закрыта; 3 – Чешуйки надрыва, выступающие кончики игл; 4 , Первое удлинение хвои примерно до двойной длины бутона; 5 , Хвоя первая, бутоны стали малярной кистью; 6 , Удлинение побега, базальная хвоя еще не распространилась; 7 , Дифференциация побега, распространение базальной хвои; 8 , Все иголки более или менее распространились, новые бутоны развиваются.
Профилирование метаболитов на основе ГХ / МС
Экстракцию образцов для анализа полярных соединений проводили в соответствии с методом, описанным Sissener et al. (2011) и Uleberg et al. (2012) с некоторыми модификациями. Объединенные образцы хвои и почек A. alba и P. abies были грубо измельчены под жидкостью N 2 , и 300 мг почек и 500 мг ткани хвои были перенесены в пробирки на 15 мл. 2000 мкл предварительно охлажденного 100% метанола с рибитом в качестве внутреннего стандарта (200 мкг / мл) добавляли к образцам и встряхивали.После обработки в течение 1 ч при 70 ° C в инкубаторе с встряхиванием в пробирки добавляли 1 мл хлороформа и, наконец, после встряхивания добавляли 1 мл H 2 O. Образцы центрифугировали 10 мин при 3000 об / мин при 4 ° C. Аликвоты из прозрачного супернатанта бутонов (800 мкл) и экстрактов игл (500 мкл) переносили в 1,5 мл круглодонные пробирки Эппендорфа и далее обрабатывали в соответствии с протоколом, описанным у Sissener et al. (2011) и хранились при -20 ° C перед ГХ / МС.
Для всех анализов использовали газовый хроматограф Varian Star 3400 CX, соединенный с масс-спектрометром Varian Saturn 3.Объем образца 1 мкл вводили с соотношением долей 25: 1. ГХ-разделение проводили на капиллярной колонке HP-5MS (30 м × 0,25 мм внутренний диаметр, толщина пленки 0,25 мкм). Температура впрыска составляла 250 ° C, а граница раздела была установлена на 250 ° C. Газ-носитель – гелий с постоянной скоростью потока 1 мл / мин. Температуру ГХ поддерживали изотермически на уровне 70 ° C в течение 5 минут, увеличивали от 70 ° C до 250 ° C со скоростью 5 ° C / мин и поддерживали при 250 ° C в течение 10 минут. Далее температуру повышали с 250 ° C до 275 ° C со скоростью 25 ° C / мин и, наконец, поддерживали при 275 ° C в течение 11 минут (время анализа: 63 мин).Источник МС был настроен на 220 ° C, и был зарегистрирован диапазон масс m / z 70–550. Все масс-спектры были получены в режиме ЭУ.
Интеграция данных ГХ / МС, нормализация (общий сигнал) и выравнивание выполнялись с использованием программного обеспечения Metalign (Lommen, 2009). Основываясь на различных количественных ионах, обнаруженные аналиты оценивались количественно на основе внутреннего стандарта рибита и, наконец, выражались в мкг / г FW. Визуализацию хроматограммы и идентификацию пиков проводили с использованием различных пакетов программного обеспечения ГХ / МС: Saturn GC / MS Workstation (Varian Analytical Instruments, Walnut Creek, CA), AMDIS (v.2,71; Национальный институт стандартов и технологий NIST, Боулдер, Колорадо) и программу с открытым исходным кодом OpenChrom Community Edition (v. 0.9.0) (Wenig and Odermatt, 2010). Предварительная идентификация соединений была достигнута с использованием базы данных метаболома Голма (GMD) (Hummel et al., 2010) и библиотеки масс-спектров NIST / EPA / NIH (NIST 05) в сочетании с собственной библиотекой индексов удерживания триметилсилилированного (TMS). ) метаболиты, обнаруженные в различных модельных видах и матрицах образцов.Всего было обнаружено 169 соединений, включая 80 метаболитов, 56 аннотированных структур и 33 неидентифицированных метаболитных метки (см. Дополнительную таблицу 1 и дополнительный рисунок 1).
Статистический анализ
Согласованные данные обрабатывались на основе комбинации самых высоких значений интенсивности ионов МС в пределах каждого окна времени удерживания (сканирование) и численности МС (обнаружение в ≥ 70% всех образцов). Всего было отфильтровано 813 меток MS и подверглось t -тестированию и двухфакторному дисперсионному анализу для оценки одиночных и комбинированных эффектов видов ( A.alba и P. abies ), ткани (бутоны и хвоя) и временные точки (9 недель). Многовариантный анализ проводился с помощью нецелевого анализа главных компонентов (PCA) с использованием программного обеспечения Minitab ® (v.17.1.0; Minitab, Ltd., Ковентри, Великобритания) (рисунки 1, 2) и иерархического кластерного анализа (HCA) с Корреляция Пирсона основана на данных, преобразованных в log2 (n) (медианная база) для отдельных меток метаболитов, с использованием программного обеспечения MultiExperiment Viewer (MeV) (версия 4.9.0) (Saeed et al., 2003) (рис. 3).Microsoft ® Excel 2010 использовался для визуализации изменений средних уровней 50 выбранных метаболитов (мкг / г FW) в ткани почек и хвои из инкубированных в теплице веточек A. alba и P. abies из 9 растений. -недельное исследование (рисунки 4, 5; дополнительный рисунок 1). Были рассчитаны корреляции (Pearson r) между оценкой раскрытия бутонов и уровнями 80 идентифицированных метаболитов, выделив метаболиты с коэффициентами корреляции r ≤ -0,5 ≤ 1 или r ≥ 0.5 ≥ 1 (рисунки 6, 7). На основе всего 80 известных и идентифицированных соединений были рассчитаны матрицы расстояния между метаболитами и визуализированы в виде тепловых карт HCA для комбинированных метаболитов почек или игл с использованием программного обеспечения MeV (дополнительный рисунок 2). Диаграммы рассеяния вулканов были построены с помощью Microsoft ® Excel 2010 на основе в общей сложности 80 идентифицированных метаболитов, обнаруженных во всех образцах ( n = 3) либо почек, либо игл из инкубированных в теплицах веточек A. alba и P . abies . Значения Log2 (кратное изменение) концентрации метаболита через 9 недель по сравнению со средним значением всех временных точек были рассчитаны и нанесены на график против -log10 (p) p -значения временных точек (дополнительный рисунок 3).
Рис. 1. Анализ главных компонентов (PCA) . 2D PCA был основан на PC1 и PC2 из средних значений концентрации ( n = 3) 36 профилей метаболитов (813 меток MS каждый). Образцы четко отделены друг от друга по породам деревьев или типам тканей.Значения ПК были получены из значений log2 (n) каждого отдельного тега MS на основе медианы для всех образцов. См. Также дополнительную таблицу 1.
Рисунок 2. Анализ главных компонентов (PCA) . 2D PCA был основан на PC1 и PC2 из значений концентраций всех повторов (всего 108 профилей метаболитов с 813 метками MS каждый). Образцы сегрегации могут быть показаны для образцов с 1-й по 9-ю неделю, которые более выражены в ткани иглы (см. Также рисунки 1, 3). Значения ПК были получены из значений log2 (n) каждого отдельного тега MS на основе медианы для всех образцов.См. Также дополнительную таблицу 1.
Рисунок 3. Иерархический кластерный анализ (HCA) на основе корреляции Пирсона, рассчитанной из средних значений концентрации ( n = 3) 36 профилей метаболитов (813 меток MS каждый) . Виды деревьев (Aa, Abies alba ; Pa, Picea abies ), тип ткани (b, почки; n, иглы) и временная точка отбора проб (номера недель от 1 до 9) представлены в столбцах, в то время как отдельные MS теги изображаются рядами. Визуализация различий на тепловой карте основана на отношении измененных уровней концентрации log2 (n) к среднему значению для каждого отдельного тега MS.Голубоватые цвета указывают на более низкие концентрации метаболитов, а красноватые цвета показывают повышенные уровни метаболитов (см. Цветовую шкалу). См. Также рисунки 1, 2 и дополнительную таблицу 1.
Рис. 4. Изменения средних уровней ( n = 3) выбранных метаболитов (мкг / г FW) в ткани почек из веточек Abies alba (-) и Picea abies (), инкубированных в теплице 9-недельный период . Планки погрешностей указывают на стандартное отклонение. См. Также дополнительный рисунок 1.
Рис. 5. Изменения средних уровней ( n = 3) выбранных метаболитов (мкг / г FW) в ткани хвои от инкубированных в теплице веточек Abies alba (-) и Picea abies (), образцы взяты из 9-недельный период . Планки погрешностей указывают на стандартное отклонение. См. Также дополнительный рисунок 1.
Результаты
Фенология бутонов
Условия теплицы ускоряют раскрытие бутонов, удлинение и созревание побегов и хвои.После 3 недели во многих пробах в апикальной зоне веточек наблюдались изменения, на которые указывало набухание почек. Значительные различия между двумя видами могут быть продемонстрированы в отношении более раннего развития почек и немного более высоких показателей раскрытия почек для P. abies по сравнению с A. alba (парный t -тест; p = 0,02824). Средние значения включены в дополнительную таблицу 1.
Метаболические сдвиги при принудительной дезакклимации
Используя нецелевой подход, 813 тегов MS, представляющих потенциальные метаболиты, были первоначально выбраны из 133 814 тегов MS из выровненного набора данных из 108 образцов анализа (дополнительная таблица 1).Качество отфильтрованных меток MS было дополнительно подтверждено на основе статистического t -тестирования и двухфакторного дисперсионного анализа, подтверждающего статистическую значимость данных по меньшей мере для одного состояния, связанного с отдельными факторами (виды, орган и ткань) или / или комбинированными эффектами. На основе этих всеобъемлющих профилей метаболитов были проведены начальные анализы PCA для описания различий между образцами в отношении видов и типов тканей (рис. 1), а также эффектов в определенные моменты времени (рис. 2). Эти два вида хвойных могут быть четко отделены друг от друга (рис. 1).Разделение образцов было еще более выраженным в отношении ткани почек и иглы, что также указывает на гораздо более сильное разделение ткани иглы A. alba и P. abies по сравнению с плотным скоплением образцов почек. Более сильное разделение временных точек (недель) в ткани иглы A. abies также можно было наблюдать при включении всех 108 повторов в анализ PCA (рис. 2). Результаты PCA определили группы выборки, как показано иерархической кластеризацией (HCA) (рисунок 3).Типы тканей были четко разделены друг от друга на два основных кластера, каждый из которых был разделен на подкластеры, относящиеся к видам хвойных. Как уже отмечалось для PCA временных точек (рис. 2), образцы ткани иглы показали почти упорядоченную кластеризацию как для A. alba , так и для P. abies , начиная с 1-й по 9-ю неделю, включая образцы почек P. abies , что указывает на сильные временные эффекты в сдвиге метаболитов.
РезультатыPCA и HCA дополнительно подтверждаются статистическим анализом с использованием тестов t и двустороннего дисперсионного анализа.Из всего 169 назначенных соединений (дополнительная таблица 1) большее количество метаболитов, показывающих значимые различия ( p ≤ 0,001), могло наблюдаться для тканевых эффектов, что указывает на дифференциальную модуляцию цикла лимонной кислоты, аминокислоты, амина, полиола. , и вторичный метаболизм. Уровни метаболитов также различались между видами, особенно те, которые участвуют во вторичном метаболизме, такие как шикимовая кислота, центральный предшественник биосинтеза ароматических аминокислот и фенольных структур, например.g., галловая кислота, эпигаллокатехин и хлорогеновая кислота, в частности, в углеводном обмене и дисахаридах. Эффекты в определенные моменты времени были не так четко выражены, но были явно затронуты центральные пути и соответствующие уровни метаболитов. Значимые вид × ткань взаимодействия были обнаружены для многих метаболитов, связанных с углеводным и вторичным метаболизмом, в то время как временная точка × ткань Эффекты были отмечены для метаболизма кислот, аминокислот и полиолов.
Изменения концентраций выбранных метаболитов за период 9 недель представлены на Рисунке 4 (почки) и Рисунке 5 (иглы).Дальнейшие результаты для 50 основных метаболитов приведены в дополнительной таблице 1. Оба показали существенные метаболические изменения к 3-й неделе дезакклимации. Независимо от типа ткани, несколько метаболитов, принадлежащих к одному и тому же пути, показали одинаковые уровни в ткани почек и хвои после обработки теплой температурой, что указывает на некоторые общие метаболические процессы в обоих типах тканей, включая метаболизм аминокислот (аспарагиновая кислота – аспарагин, изолейцин – валин и серин – глицин – треонин), метаболизм углеводов (сахароза – G6P – F6P), жирные кислоты (C16: 0 – C18: 0) и промежуточных продуктов цикла Кребса (янтарная кислота – фумаровая кислота – яблочная кислота).Эти результаты были дополнительно подтверждены взаимосвязями путей в общей сложности 80 идентифицированных метаболитов в ткани почек или хвои обоих видов хвойных, при визуализации на корреляционных тепловых картах (дополнительный рисунок 2). Сгруппированная матрица расстояний между метаболитами и метаболитами дает исчерпывающее представление о различиях и сходствах в регуляции метаболизма и подчеркивает взаимосвязь метаболических модулей и отдельных путей (например, жирных кислот и углеводов). Напротив, некоторые соединения, которые рассматриваются не как промежуточные продукты биосинтеза, а как конечные метаболиты (например,g., вторичные метаболиты) и / или известные совместимые растворенные вещества (например, пролин) продемонстрировали характерные закономерности накопления или разложения на протяжении всего тепличного исследования.
В то время как концентрация некоторых метаболитов в почках снизилась (например, пролина, глицерина и катехина), многие метаболиты продемонстрировали отчетливо повышенные уровни, связанные с развитием, ростом и фотосинтезом бутонов (например, лимонная и яблочная кислоты, пинитол, ксилоза, рибоза, мелибиоза, аспарагиновая кислота и аспарагин) (рис. 4).При сравнении видов A. alba , как правило, демонстрировал более раннее начало метаболической модуляции и большие изменения концентрации по сравнению с P. abies . Это особенно верно для многих пентоз и дисахаридов, фосфатов, некоторых вторичных метаболитов, кислот цикла Кребса и аминокислот (пироглутаминовая кислота и ГАМК). Также наблюдались видовые различия в отношении общего уровня метаболитов с высоким содержанием на основе средних значений (дополнительная таблица 1 и дополнительный рисунок 1), включая сахара, такие как ксилоза, рибоза, фруктоза и сахароза, спирты (пинитол и мио -инозит) , кислоты (хинная кислота) и родственные катехину соединения.Инициирование метаболизма и основные биохимические изменения в иглах A. alba и P. abies , по-видимому, были отложены по сравнению с тканью почек (Рисунок 5 и Дополнительный Рисунок 1). Однако большинство аминокислот показали очевидную реакцию на воздействие теплых условий (например, аланин, валин, глутаминовая кислота и аспарагиновая кислота), часто показывая характерные переходные пики концентрации. Значительное снижение уровней янтарной, фумаровой и яблочной кислоты в начале обработки в теплице сопровождалось увеличением к концу 9-недельного периода.Пентозы и гексозы также увеличивались в иглах обоих видов. Несколько основных метаболитов игл P. abies (дополнительная таблица 1 и дополнительная фигура 1) показали, как правило, более высокие уровни концентрации (средние значения), включая аминокислоты, отдельные сахара (например, мальтозу и галактозу), кислоты (хинная кислота, шикимовая кислота). , и яблочная кислота) и вторичные структуры (например, хлорогеновая кислота), и, таким образом, подчеркивают биохимические различия между видами. Более того, метаболические сдвиги при принудительной деакклимации часто демонстрируют обратные тенденции, примером которых являются дисахариды (мальтоза, мелибиоза, сахароза, G6P и F6P) и вторичный метаболизм (например.g., шикимовая кислота и катехин) в A. alba и P. abies .
Корреляция метаболизма с развитием почек
В почках A. alba несколько углеводов и родственных метаболитов (сахарные спирты, кислоты и фосфаты) показали положительную корреляцию от средней до сильной между измеренной за неделю концентрацией и соответствующей оценкой раскрытия почки (Рисунок 6), в то время как в бутонах Норвегии яблочная кислота ели, лимонная кислота, пинитол, ксилоза и рибоза тесно коррелировали с показателем раскрытия бутонов.Некоторые вторичные соединения показали положительную корреляцию с показателем раскрытия бутонов у обоих видов, но эпикатехин флаван-3-ол ( A. alba ) и катехин ( P. abies ) коррелировали отрицательно. Только глутаминовая кислота и аспарагин показали повышение уровня в соответствии с развитием почек у A. alba , тогда как 5 аминокислот имели положительную корреляцию в почках P. abies . Общей чертой является явно отрицательная ассоциация пролина, тогда как биосинтез винной кислоты сильно связан с открытием почек у обоих видов.
По сравнению с бутонами, иглы обоих видов показали относительно более сильную положительную корреляцию связанных с сахаром метаболитов (ксилозы, рибозы, фруктозы и глюкозы) с показателем раскрытия бутонов (Рисунок 7). Очевидные положительные отношения наблюдались для мальтозы и G6P у A. alba , в отличие от P. abies . Как уже указывалось для ткани почек, относительно сильная взаимосвязь метаболитов цикла Кребса была также обнаружена в иглах P. abies , на что указывает положительная корреляция для яблочной и фумаровой кислот и отрицательная корреляция для янтарной кислоты.От умеренной до сильной отрицательной корреляции концентраций метаболитов с количеством почек в ткани хвои была указана для сахарного спирта myo -инозитола у обоих видов, в то время как аспарагиновая кислота и хлорогеновая кислота были связаны положительно. Более того, графики Volcano кратных изменений метаболитов, построенные против p -значений временных точек (недель), были построены для выявления тех соединений со значительными изменениями уровней концентрации в конце периода деакклимации на 9 неделе (дополнительный рисунок 3).Дифференциальная регуляция биосинтеза сахарных кислот (галактоновая кислота, галактуроновая кислота и глюкуроновая кислота) и сахаров (галактоза) также указывает на модуляцию метаболизма аскорбата и альдарата в отвердевших растительных тканях. Результаты дополнительно подчеркивают характерные сдвиги в отдельных путях, выделяя как отдельные соединения с известной или потенциальной функцией осмолита, такие как пролин (все типы тканей), аспарагин (иглы), изолейцин и валин (иглы P. abies ) и путресцин (почки). А.alba ). Последнее соединение превращается в 4-аминомасляную кислоту (ГАМК) и последовательно связывает метаболизм глутамата и цикл Кребса через шунт ГАМК (см. Дополнительный рисунок 1 и таблицу 1), что указывает на взаимосвязь метаболических модулей в физиологических процессах, вызванных температурой.
Обсуждение
Фенология взрыва почек
Глобальное потепление может повлиять на рост и развитие растений, включая такие фенологические явления, как распускание почек, опадение, старение и цветение.На эти события может повлиять повышение глобальных температур, приводящее к изменению ритма развития лесных деревьев в сторону раннего начала и прекращения распускания почек (Ceulemans and Mousseau, 1994; Morin et al., 2009). В настоящем исследовании образцы роста почек наблюдались в течение 9 недель при принудительной деакклимации и распускании почек. Хотя распускание почек на отделившихся веточках было неполным через 9 недель, оно выглядело нормальным на стадии набухания почек (стадия 3) и на ранних стадиях распускания почек.Однако представленные здесь результаты необходимо учитывать в отношении экспериментальной установки (контролируемая среда при постоянной температуре и неизменных условиях дневного света) и потенциально нарушенном физиологическом функционировании (взаимоотношения источник-сток) из-за отделения побегов. В аналогичных исследованиях с использованием принудительной деакклимации с фотопериодами длинного дня растения показали усиленный рост при промывании хвои (Jach and Ceulemans, 1999) и более раннее распускание почек (Riikonen et al., 2012). Повышенные температуры сосны обыкновенной привели к более раннему отвердеванию и потенциальному повреждению морозом (Hänninen et al., 1993; Репо и др., 2006). В зависимости от продолжительности воздействия высоких температур и вида хвойных растений степень повреждения морозом может сильно различаться (Gooch et al., 2009). У нехвойных пород деревьев, таких как камедь ( Eucalyptus pauciflora ), относительно небольшие изменения дневных температур в мягкую погоду ранней весной в сочетании с повышенными уровнями CO 2 могут быть достаточными, чтобы вызвать повреждение листьев морозом ( Loveys et al., 2006). Любое дальнейшее повышение температуры уравновесит вегетативную продуктивность снежной камеди, а также снизит преимущества высокой фотосинтетической активности (Lutze et al., 1998).
Изменения в пулах метаболитов, связанные с развитием зародышей и побегов
На основе профилирования ГХ / МС и оценки фрагментов МС было обнаружено в общей сложности 813 меток МС во всех образцах, из которых было обнаружено 169 назначенных метаболитов, многие из которых, как известно, участвуют в различных метаболических процессах у хвойных деревьев во время деакклимации. Учитывая полярность экстракционного растворителя, обнаружение жирных кислот было естественным низким, то есть диапазон обнаруженных и идентифицированных соединений охватывал в основном полярные метаболиты, обычно содержащие несколько различных функциональных групп, присоединенных к молекуле.Другим фактором, касающимся ограничения обнаружения соединений, были применяемые настройки ГХ / МС в сочетании с условиями экстракции, исключающие как стерины, так и трисахариды. Однако последние функционально участвуют в процессах акклиматизации к холоду и приобретают морозостойкость, как, например, в случае раффинозы (Rohloff et al., 2012; Davik et al., 2013), и легко катаболизируются до дисахаридов и гексоз во время деакклимации (Strimbeck и др., 2008). Предварительная идентификация соединений на основе поиска в базе данных MS с использованием собственной MS-библиотеки метаболитов TMS и индексов удерживания привела к относительно высокому количеству идентифицированных метаболитов 80, соединений с известным молекулярным классом 55 и неизвестных структур 34 (всего 169 назначенные метаболиты).Однако, учитывая первоначальный выбор 813 меток MS и режим фильтрации, несколько потенциальных метаболитов могли быть упущены. В зависимости от чувствительности прибора (TOF против ионной ловушки против квадруполя) и настроек, исследования профилей метаболитов у различных видов хвойных с использованием ГХ / МС дали аналогичные результаты с общим количеством назначенных метаболитов в диапазоне от 44 до 115 идентифицированных соединений (например, Businge et al. al., 2012 [44 метаболита]; Eckert et al., 2012 [86]; Shiryaeva et al., 2012 [115]; Angelcheva et al., 2014 [68]; Lee et al., 2014 [105]), тем самым подчеркивая качество детекции ГХ / МС и применимость метода для регистрации изменений в пулах метаболитов.
В нашем исследовании мы впервые представляем углубленный анализ регуляции метаболизма во время отвердевания и раскрытия почек в тканях хвойных почек и хвои. Всего 80 идентифицированных соединений дают подробный обзор моделей метаболитов и изменений во время принудительной деакклимации. Судьба и закономерности накопления и деградации углеводов могут быть объяснены физиологически, выявив возможное выделение и распределение различных метаболитов в хвое и почках A.alba и P. abies . В целом, изменения в уровнях углеводов и других растворенных веществ связаны с сезонными колебаниями акклиматизации и дезакклимации древесных видов (Cox and Stushnoff, 2001; Strimbeck et al., 2008).
К сожалению, настоящая экспериментальная установка ограничивает возможность экспорта сахара, потому что веточки, отделенные от родительского растения, отбирались и выращивались в теплице без возможности перемещения фотосинтатов и метаболитов в корни.Поскольку в прошлом году хвоя молодых побегов была полностью дифференцированной и способной к фотосинтезу, фотосинтетический эффект у хвои был потенциально очень высоким. Фотосинтаты в новообразованных иголках и почках являются основным источником углерода для развития побегов, и общее увеличение весеннего сахара хорошо задокументировано (Fischer and Höll, 1991). Когда иглы достигают половины своего окончательного роста и могут легко поставлять другие продукты фотосинтеза, они перемещаются к корням (Ericsson, 1978).Однако в нашем эксперименте транслокация тормозилась из-за отсутствия корневой системы.
Оказалось, что изменения в уровнях концентрации отдельных углеводов больше влияют на хвою, чем на почки. Эти изменения сахаров, которые могут действовать как осмолиты, могут усиливать движение воды к развивающейся почке (De Faÿ et al., 2000). В древесине лиственных деревьев количество продуктов фотосинтеза уменьшается к финальной стадии распускания почек с последующим промыванием новых побегов (Kozlowski, Keller, 1966).Однако у хвойных деревьев развитие новых побегов не зависит от запаса, накопленного от хвои предыдущего года, т. Е. Побеги могут легко поддерживать себя фотосинтетическим материалом. К концу развития хвои скорость фотосинтеза увеличивается вместе с повышенной мобилизацией метаболитов в хвое и почках, и аналогично уровни метаболитов в стеблях повышаются в несколько раз в конце лета (Hansen and Beck, 1994). Улучшение фотосинтеза в зависимости от температуры хорошо задокументировано (Corcuera et al., 2005), из которого следует, что температура увеличивает использование запасов углерода в растениях. Таким образом, настоящее исследование демонстрирует аналогичные эффекты ранней весенней температуры, когда почки и хвоя демонстрируют четко разграниченные различия в ответ на температуру, полученную в теплице.
Хвойные деревья выработали несколько механизмов, позволяющих справляться с неблагоприятными климатическими изменениями. У Pinus sylvestris низкая температура приводит к снижению фотосинтетической активности и, следовательно, к снижению первичного роста (Ensminger et al., 2006). В природе дезакклимация начинается медленно и постепенно достигает максимума с наступлением весны. После перерыва в покое и обезвоживания растений снова вернуться к фазе акклиматизации невозможно. Этот эффект называется пассивной и медленной дезакклимацией; следовательно, хвойные деревья очень медленно достигают фотосинтетической активности. В нашем исследовании в контролируемых условиях окружающей среды увеличение общих продуктов фотосинтеза наблюдалось на ранних стадиях вместе с инициированием метаболизма.После того, как акклиматизированные к холоду растения подвергаются воздействию повышенных и «неестественно» высоких температур, дезакклимация начинается в течение 2 или 3 часов (Chen and Li, 1980), и зимостойкость может быть потеряна в течение одного или нескольких дней, как обсуждали Kalberer et al. (2006). Прямое применение постоянных повышенных температур (22 ° C) в тепличных условиях привело к высокой метаболической активности, т.е. фотосинтез считался эффективным, на что указывало устойчивое увеличение сахаров после начального падения концентрации (например,g., фруктоза и глюкоза) (дополнительный рисунок 1), что также подтверждается положительной корреляцией метаболитов (рисунки 6, 7). В природе мы скорее наблюдаем противоположное явление с постепенным повышением температуры, инициированием клеточного дыхания и разложением запасных сахаров и осмолитов. В то же время фотосинтез все еще находится на низком уровне, не происходит накопления достаточного количества ассимилятов для повышения уровня сахара, поэтому ожидается, что уровни фруктозы и глюкозы упадут при отвердевании акклиматизированной к холоду растительной ткани в естественных условиях окружающей среды (Strimbeck et al., 2008). Напротив, почки хвойных растений в нашем исследовании не демонстрировали таких закономерностей не в последнюю очередь из-за необходимости установить фотосинтетические функции и из-за быстро метаболизируемых ассимилятов.
Наблюдаемые паттерны деградации метаболитов могут быть связаны с потребностью в компонентах для синтеза материала клеточной стенки и другими энергетическими потребностями в развивающихся иглах (Hansen and Beck, 1990). A. alba , который естественным образом адаптирован к более теплому климату по сравнению с елью обыкновенной, также продемонстрировал более раннее начало повышения уровней связанных с энергией метаболитов, таких как фосфорная кислота, и ключевых промежуточных продуктов в гликолизе / глюконеогенезе, таких как F6P и G6P.Эти наблюдения дополнительно подтверждаются общим увеличением уровней кислот цикла Кребса в почках (дополнительный рисунок 1), то есть лимонной и яблочной кислоты у обоих видов, а также янтарной и фумаровой кислот (у P. abies ). Положительная корреляция кислот цикла Кребса с показателем раскрытия почек в бутонах и иглах P. abies (рисунки 6B, 7B) продемонстрировала ключевую роль цикла Кребса в отношении фотосинтеза, центрального метаболизма и производства энергии.
Сравнение метаболизма почек и хвои A.alba и P. abies с использованием корреляции Пирсона (данные не представлены) выявили, что изменения уровней концентрации высокоразвитых центральных метаболитов часто демонстрируют сходные тенденции у обоих видов хвойных (дополнительный рисунок 1). Важные соединения включали: глюкозу (иглы: r = 0,54), фруктозу (бутоны: 0,69) и ксилозу (бутоны: 0,60; иглы: 0,58), кислоты цикла Кребса, лимонную кислоту (бутоны: 0,75) и яблочную кислоту (иглы: 0,63), и соединения, связанные с метаболическими путями, такие как хинная кислота (почки: 0.75) и пинитол (бутоны: 0,73). Напротив, многие метаболиты демонстрировали обратные модели модуляции между видами, включая дисахариды, аминокислоты и вторичные соединения, то есть как время, так и степень модуляции пулов метаболитов сильно различались между центральноевропейскими видами A. alba и P. abies происходит из Северной Европы.
Модуляция пулов метаболитов, связанных с накопленными осмолитами
Превращение крахмала в растворимые сахара и увеличение концентрации углеводов обычны для растений во время акклиматизации и адаптации к стрессу замораживания (Sakai and Larcher, 1987).Тем не менее, накопление совместимых растворенных веществ с функцией криопротектора не ограничивается сахарами и их олигомерами, а скорее связано с антиоксидантной активностью и модуляцией других растворенных веществ и полимерных структур, таких как аминокислоты и амины, органические кислоты, полиолы, фосфолипиды, и белки (Ruelland et al., 2009). Было показано, что накопление / разложение сахаров во время акклиматизации / обезвоживания различается между хвойными деревьями, произрастающими в северных и умеренных регионах (Strimbeck et al., 2007, 2008). Это означает, что прямое сравнение различных концентраций метаболитов между исследуемыми видами в нашем исследовании может вводить в заблуждение, и следует скорее сосредоточиться на начале и времени метаболических изменений в ткани почек и хвои. Точно так же, как акклиматизация растений к холоду достигается за счет согласования экспрессии генов и лежащих в их основе регуляторных и метаболических сетей, как впервые показано на модели растения Arabidopsis thaliana (например, Cook et al., 2004; Kaplan et al., 2004, 2007), дезакклимация и потенциальная реакклимация аналогичным образом характеризуются согласованными регуляторными паттернами (Byun et al., 2014). Очевидно, что динамический характер обезвоживания растений связан не только с генотипическими различиями, но и напрямую определяется условиями окружающей среды во время фазы акклиматизации растений (Kalberer et al., 2006), обычно называемой воспоминанием о холодовом стрессе (Byun et al., 2014). Предполагая равные условия акклиматизации к холоду, которым родительские виды и генотипы в нашем исследовании подвергались зимой, и из-за контролируемых тепличных условий, в которых выращивались образцы побегов, дифференциальную регуляцию метаболизма следует рассматривать в контексте индивидуальных генотипических вариаций ( стандартное отклонение), но, что наиболее важно, в отношении видовых различий между A.alba и P. abies .
Хвойные растения при теплой температуре не могут полностью деакклиматизировать свои ткани ниже минимального уровня выносливости до тех пор, пока состояние покоя не будет нарушено, но после завершения любое дальнейшее повышение температуры может напрямую повлиять и увеличить рост растений с последующей регидратацией (Kalberer et al. ., 2006). В естественных условиях необходимо медленно дезакклимировать растения, чтобы предотвратить повреждение тканей морозом (Saxe et al., 2001). Принудительная деакклимация инициировала быструю деградацию и / или модуляцию накопленных осмолитов, потенциально связанных с морозостойкостью.Одним из наиболее известных метаболитов, которые, как известно, участвуют в механизмах толерантности к холоду, является аминокислота пролин (Cook et al., 2004). Как наблюдали в ткани почек обоих видов хвойных, уровни пролина быстро снижались с 300-500 мкг / г FW до 30-70 мкг / г FW после 9-недельной обработки теплой температурой (корреляция Пирсона: r = 0,88). Это соответствует недавним открытиям Lee et al. (2014), которые сообщили о резком повышении уровня пролина в верхушках побегов ели европейской во время фазы акклиматизации к холоду.Напротив, мы не обнаружили снижения других осмолитических аминокислот, таких как глутаминовая кислота, аспарагин (Lee et al., 2014) и валин (Angelcheva et al., 2014). Мы обнаружили корреляцию между видами в одной или обеих тканях в различных аминокислотах (дополнительный рисунок 1): валин (почки: r = 0,82; иглы: 0,69), серин (почки: 0,62), треонин (почки: 0,55; иглы: 0,66), аспарагиновая кислота (бутоны: 0,52; хвоя: 0,81) и аспарагин (бутоны: 0,76). P. abies показал более высокие уровни концентрации этих соединений, и, кроме того, увеличение количества аминокислот в хвое ели европейской произошло в более ранний момент времени, включая аланин, пролин и пироглутаминовую кислоту, что указывает на более выраженные реакции метаболизма азота у этого вида.Подобные дифференциальные ответы в иглах наблюдались также для нескольких пентоз и гексоз, несмотря на тесную корреляцию уровней метаболитов между видами, как уже обсуждалось, включая дисахариды, такие как сахароза, мальтоза и мелибиоза. Как правило, повышение уровня последнего метаболита указывает на устойчивую деградацию (предполагаемых) высоких запасов рафинозы (Cox and Stushnoff, 2001; Strimbeck et al., 2008; Lee et al., 2014) и последовательную мобилизацию углеводных резервов, которая больше не требуется для сохранять морозостойкость.
Было обнаружено, что пулы нескольких небольших молекул с известной или предполагаемой функцией осмосопротектора быстро уменьшаются во время принудительной деакклимации в ткани иглы, включая глицерин, яблочную кислоту и янтарную кислоту. Эти результаты согласуются с более ранними исследованиями, указывающими на повышение уровня малата и сукцината во время акклимации к холоду у хвойных пород (Dauwe et al., 2012; Angelcheva et al., 2014; Lee et al., 2014), которые позже могут деградировать во время фаза отверждения. Однако обнаруженные уровни соединений были сравнительно низкими (за исключением яблочной кислоты), поэтому потенциальная роль метаболитов в функционировании осмолита остается неясной.В какой степени вторичные метаболиты, такие как простые фенольные структуры и полифенолы, непосредственно и функционально участвуют в процессах акклимации / деакклимации растений к холоду, помимо их антиоксидантной активности, все еще остается предметом обсуждения. Несколько авторов сообщили о повышении уровня флавоноидов в тканях листьев хвойных пород осенью / в начале зимы, таких как катехин (Dauwe et al., 2012). Наши результаты показывают исходное снижение уровней катехина и эпикатехина (только A.alba ) в бутонах, соответствуют исследованиям Pukacki и Kaminìska-Rożek (2013), которые отслеживали снижение общего количества флавоноидов у P. abies во время деакклимации, и соответствуют другим отчетам, использующим подходы к профилированию метаболитов для характеристики моделей накопления различные соединения (например, катехин) при акклиматизации к холоду у P. obovata (Angelcheva et al., 2014), P. sitchensis (Dauwe et al., 2012) и P. abies (Lee et al. , 2014).Кроме того, наши данные свидетельствуют о тесной взаимосвязи различных модулей вторичного метаболизма, включая такие предшественники, как шикимовая кислота и катехины (почки) и эпигаллокатехин с галловой кислотой (иглы) (дополнительный рисунок 2). Недавние функциональные исследования, интегрированные молекулярные подходы с использованием передовых биоинформатических инструментов пролили свет на роль вторичного метаболизма в толерантности растений к холоду (например, Usadel et al., 2008; Doerfler et al., 2013) и, надеюсь, также будут касаться происходящих биохимических процессов. на этапе дезаклимации в ближайшем будущем.
Заключение
Изменение климата и глобальное потепление, как правило, приводят к повышению средних температур и риску наступления теплых периодов, что приводит к более раннему началу роста, распусканию почек и потенциальному повреждению морозом древесных пород бореальных лесов. В настоящем исследовании мы исследовали фенологию почек на побегах двух видов хвойных, Abies alba и Picea abies , во время принудительной дезакклимации в тепличных условиях. ГХ / МС профилирование метаболитов ткани почек и хвои было использовано здесь впервые в комплексном подходе для измерения изменений в пулах метаболитов в течение 9 недель подряд.На основе нецелевого многомерного анализа (PCA и HCA) и обнаружения 169 назначенных метаболитов, включающих 80 идентифицированных соединений, мы смогли охарактеризовать основные изменения в уровнях углеводов, аминокислот, органических кислот и вторичных метаболитов. Корреляционный анализ выявил взаимосвязь метаболитов, связанных с метаболическими путями, и взаимосвязь метаболических модулей. Кроме того, на основе корреляции фенологических данных и данных о метаболитах, а также анализа кратных изменений метаболитов, мы смогли точно определить те метаболиты и структурные классы, которые были в значительной степени связаны с распусканием почек и инициированием роста.Мы смогли четко продемонстрировать дифференциальную регуляцию метаболизма в почках и листьях хвои обоих видов хвойных. Различия между видами были сильно выражены, хотя уровни нескольких метаболитов из разных классов соединений демонстрировали сходные модели модуляции, такие как аминокислоты, кислоты цикла Кребса и сахара. Однако начало и степень изменений указали на различия между видами хвойных, адаптированных к бореальному климату ( P. abies ) и A. alba , происходящим из зон умеренного климата.В заключение, настоящие результаты пролили свет на эффекты вызванных температурой изменений в метаболизме растений в первоначально спящей ткани и будут дополнительно поддерживать дискуссии о влиянии прогнозируемого глобального потепления на морозостойкость, фенологические явления и биохимические изменения деревьев во время дезакклимация.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: http://www.frontiersin.org/Journal/10.3389/fpls.2014.00706/abstract
Список литературы
Ангелчева, Л., Мишра, Ю., Антти, Х., Кьельсен, Т. Д., Функ, К., Стримбек, Г. Р. и др. (2014). Метаболомический анализ экстремальной морозостойкости ели сибирской ( Picea obovata ). Новый Фитол . 204, 545–555. DOI: 10.1111 / Nph.12950
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бреймейер, А.И., Берг Б., Гауэр С. Т. и Джонсон Д. В. (1996). «Углеродный бюджет: хвойные леса умеренного пояса», в Global Change: Effects on Coniferous Forests and Grasslands , ред. А. И. Бреймейер, Д. О. Холл, Дж. М. Мелилло и Г. И. Агрен (Чичестер: John Wiley & Sons, Inc.), 41–68 .
Бусинге, Э., Бракманн, К., Мориц, Т., и Эгертсдоттер, У. (2012). Профилирование метаболитов выявляет четкие метаболические изменения во время соматического развития зародыша ели европейской ( Picea abies ). Tree Physiol . 32, 232–244. DOI: 10.1093 / treephys / tpr142
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Бьюн, Й.-Дж., Ку, М.-Й., Джу, Х.-Дж., Ха-Ли, Ю.-М., и Ли, Д.-Х. (2014). Сравнительный анализ экспрессии генов в условиях холодовой акклимации, дезакклимации и реакклимации у арабидопсиса . Physiol. Завод . 152, 256–274. DOI: 10.1111 / ppl.12163
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Каннелл, М.Г. Р. (1990). Моделирование фенологии деревьев. Silva Carelica 15, 11–27.
Google Scholar
Ceulemans, R., and Mousseau, M. (1994). Обзор Тэнсли № 71: воздействие повышенного содержания CO в атмосфере 2 на древесные растения. Новый Фитол . 127, 425–446. DOI: 10.1111 / j.1469-8137.1994.tb03961.x
CrossRef Полный текст | Google Scholar
Кук, Д., Фаулер, С., Файн, О., и Томашоу, М. Ф. (2004). Видная роль пути холодовой реакции CBF в настройке низкотемпературного метаболома Arabidopsis . Proc. Natl. Акад. Sci. США . 101, 15243–15248. DOI: 10.1073 / pnas.0406069101
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Коркуера, Л., Моралес, Ф., Абадия, А., и Гиль-Пелегрин, Э. (2005). Сезонные изменения фотосинтеза и фотозащиты у Quercus ilex subsp. Балтийский лесной массив, расположенный на его верхнем высотном пределе на Пиренейском полуострове. Tree Physiol . 25, 599–608. DOI: 10,1093 / treephys / 25.5,599
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кокс, С. Э., и Стушнофф, К. (2001). Температурные сдвиги в содержании растворимых углеводов в период покоя и акклиматизации к холоду у Populus tremuloides . Банка. J. Forest Res . 31, 730–737. DOI: 10.1139 / x00-206
CrossRef Полный текст | Google Scholar
Даув, Р., Холлидей, Дж. А., Эйткен, С. Н., и Мэнсфилд, С. Д. (2012). Динамика метаболизма во время осенней акклиматизации к холоду внутри и среди популяций ели ситкинской ( Picea sitchensis ). Новый Фитол . 194, 192–205. DOI: 10.1111 / j.1469-8137.2011.04027.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Davik, J., From, B., Koehler, G., Torp, T., Rohloff, J., Eidem, P., et al. (2013). Уровни дегидрина, алкогольдегидрогеназы и центральных метаболитов связаны с холодоустойчивостью диплоидной клубники ( Fragaria spp.). Planta 237, 265–277. DOI: 10.1007 / s00425-012-1771-2
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
De Faÿ, E., Вашер, В., и над Гумберт, Ф. (2000). Связанные с водой явления в озимых почках и ветках Picea abies L. (Karst.) До распускания почек: биологическое, гистологическое и ЯМР исследование . Ann. Бот . 86, 1097–1107. DOI: 10.1006 / anbo.2000.1276
CrossRef Полный текст
Диксон, Р.К., Соломон, А.М., Браун, С., Хоутон, Р.А., Трексье, М.С., и Вишневски, Дж. (1994). Запасы углерода и потоки глобальных лесных экосистем. Наука 265, 185–190.DOI: 10.1126 / science.263.5144.185
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Добровольская Д. (1998). Структура естественного возобновления пихты пихты ( Abies alba Mill.) В заповеднике «Ята» в Польше. Forest Ecol. Манаг . 110, 237–247. DOI: 10.1016 / S0378-1127 (98) 00286-2
CrossRef Полный текст | Google Scholar
Doerfler, H., Lyon, D., Nägele, T., Sun, X., Fragner, L., Hadacek, F., et al. (2013).Причинная связь по Грейнджеру в интегрированных данных метаболомики ГХ-МС и ЖХ-МС раскрывает границу первичного и вторичного метаболизма. Метаболомика 9, 564–574. DOI: 10.1007 / s11306-012-0470-0
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Eckert, A. J., Wegrzyn, J. L., Cumbie, W. P., Goldfarb, B., Huber, D. A., Tolstikov, V., et al. (2012). Генетика ассоциации метаболома сосны лоблольной ( Pinus taeda , Pinaceae). Новый Фитол .193, 890–902. DOI: 10.1111 / j.1469-8137.2011.03976.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Эггер, Б., Эйниг, В., Склерет, А., Валленда, Т., Магель, Э., Лоу, А. и др. (1996). Углеводный обмен в однолетней и двухлетней хвое ели и стеблевые углеводы от трех месяцев до и до трех месяцев после распускания почек. Physiol. Завод . 96, 91–100. DOI: 10.1111 / j.1399-3054.1996.tb00188.x
CrossRef Полный текст | Google Scholar
Эрикссон, А.(1978). Сезонные изменения транслокации 14С из разных возрастных классов хвои на 20-летних деревьях сосны обыкновенной ( Pinus silvestris ). Physiol. Завод . 43, 351–358. DOI: 10.1111 / j.1399-3054.1978.tb01593.x
CrossRef Полный текст | Google Scholar
Файн, О., Копка, Дж., Третуэй, Р. Н., и Виллмитцер, Л. (2000). Идентификация необычных метаболитов растений на основе расчета элементного состава с помощью газовой хроматографии и квадрупольной масс-спектрометрии. Анал. Chem . 72, 3573–3580. DOI: 10.1021 / ac9i
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Фишер К. и Хёлль В. (1991). Пищевые запасы сосны обыкновенной ( Pinus sylvestris L.). I. Сезонные изменения углеводных и жировых запасов хвои сосны. Деревья 5, 187–195. DOI: 10.1007 / BF00227524
CrossRef Полный текст | Google Scholar
Гуч, Н. Дж., Нзоку, П., и Крегг, Б.М. (2009). Влияние пребывания в помещении на морозостойкость и физиологию контейнерных елок. Horttechnology 19, 72–77.
Google Scholar
Ханнерц, М. (1999). Оценка температурных моделей для прогнозирования распускания почек у ели европейской. Банка. J. Forest Res . 29, 9–19. DOI: 10.1139 / x98-175
CrossRef Полный текст | Google Scholar
Ханнинен, Х., Келломаки, С., Лайтинен, К., Паяри, Б., и Репо, Т. (1993). Влияние повышенной зимней температуры на начало высотного роста сосны обыкновенной: полевые испытания фенологической модели. Сильва Фенн . 4, 251–257. DOI: 10.14214 / sf.a15679
CrossRef Полный текст
Хансен, Дж., И Бек, Э. (1990). Судьба и путь продуктов ассимиляции в стволе 8-летней сосны обыкновенной ( Pinus sylvestris ). Деревья 4, 16–21. DOI: 10.1007 / BF00226235
CrossRef Полный текст | Google Scholar
Хансен, Дж., И Бек, Э. (1994). Сезонные изменения использования и оборота продуктов ассимиляции у 8-летней сосны обыкновенной ( Pinus sylvestris L.) деревья. Деревья 8, 172–182. DOI: 10.1007 / BF00196844
CrossRef Полный текст | Google Scholar
Хайде, О. М. (1993). Ответы на продолжительность светового дня и тепловое время распускания почек во время выхода из покоя у некоторых северных лиственных деревьев. Physiol. Завод . 88, 531–540. DOI: 10.1111 / j.1399-3054.1993.tb01368.x
CrossRef Полный текст | Google Scholar
Хаммель, Дж., Стремель, Н., Селбиг, Дж., Вальтер, Д., и Копка, Дж. (2010). Дерево решений поддерживало предсказание субструктуры метаболитов на основе профилей ГХ-МС. Метаболомика 6, 322–333. DOI: 10.1007 / s11306-010-0198-7
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
IPCC. (2007). «Изменение климата 2007: основы физических наук» в Вклад Рабочей группы I в Четвертый оценочный отчет Межправительственной группы экспертов по изменению климата (Нью-Йорк, Нью-Йорк: издательство Кембриджского университета).
Як, М. Э., и Сеулеманс, Р. (1999). Влияние повышенного содержания CO в атмосфере 2 на фенологию, рост и структуру кроны сеянцев сосны обыкновенной ( Pinus sylvestris ) после двух лет воздействия в поле. Tree Physiol . 19, 289–300. DOI: 10.1093 / treephys / 19.4-5.289
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Кальберер, С. Р., Вишневски, М., и Арора, Р. (2006). Деакклимация и реакклимация морозостойких растений: современное понимание и новые концепции. Завод Sci . 171, 3–16. DOI: 10.1016 / j.plantsci.2006.02.013
CrossRef Полный текст | Google Scholar
Каплан, Ф., Копка, Дж., Дейл, В.H., Zhao, W., Schiller, K.C., Gatzke, N., et al. (2004). Изучение метаболома температурного стресса Arabidopsis . Физиология растений . 136, 4159–4168. DOI: 10.1104 / стр.104.052142
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Каплан Ф., Копка Дж., Сунг Д. Ю., Чжао В., Попп М., Порат Р. и др. (2007). Профилирование транскриптов и метаболитов во время акклиматизации Arabidopsis к холоду выявляет сложную взаимосвязь между экспрессией регулируемых холодом генов и модификациями содержания метаболитов. Завод J . 50, 967–981. DOI: 10.1111 / j.1365-313X.2007.03100.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Крутч П. (1973). Развитие почек у ели обыкновенной. Внутренний отчет, IUFRO S2.02.11 . Вена: Международный союз лесных исследовательских организаций.
Ларчер, В. (2003). Физиологическая экология растений – экофизиология и стресс-физиология функциональных групп . Берлин; Нью-Йорк: Спрингер.DOI: 10.1007 / 978-3-662-05214-3
CrossRef Полный текст | Google Scholar
Лаванда, Д. П., и Силим, С. Н. (1987). Роль регуляторов роста растений в покое лесных деревьев. Регулятор роста растений . 6, 171–191. DOI: 10.1007 / BF00043954
CrossRef Полный текст | Google Scholar
Ли, Ю. К., Александер, Д., Вульф, Дж., И Олсен, Дж. Э. (2014). Изменения в профилях метаболитов в верхушках побегов ели европейской во время короткого дня индуцировали развитие зимних почек, а длинные – индуцированное распускание почек. Метаболомика 10, 842–858. DOI: 10.1007 / s11306-014-0646-x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лисек, Дж. Н., Шауэр, Н., Копка, Дж., Уиллмитцер, Л., и Ферни, А. Р. (2006). Газовая хроматография, масс-спектрометрия, профилирование метаболитов в растениях. Nat. Протокол . 1, 387–396. DOI: 10.1038 / nprot.2006.59
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Лутце, Дж.Л., Роден, Дж. С., Холли, К. Дж., Вулф, Дж., Эгертон, Дж. Дж. Г. и Болл, М. С. (1998). Повышенная атмосферная [CO 2 ] способствует повреждению саженцев вечнозеленых деревьев морозом. Среда растительных клеток . 21, 631–635. DOI: 10.1046 / j.1365-3040.1998.00296.x
CrossRef Полный текст | Google Scholar
Моэн, А. (1999). Национальный атлас Норвегии: растительность . Хёнефосс: Норвежское картографическое управление.
Morin, X., Lechowicz, M. J., Augspurger, C., O’Keefe, J., Винер Д., Чуин И. (2009). Фенология листьев у 22 видов деревьев Северной Америки в 21 веке. Glob. Сменить Biol . 15, 961–975. DOI: 10.1111 / j.1365-2486.2008.01735.x
CrossRef Полный текст | Google Scholar
Мюррей М. Б., Каннелл М. Г. Р. и Смит Р. И. (1989). Дата появления почек у пятнадцати видов деревьев в Великобритании после потепления климата. J. Appl. Экол . 26, 693–700. DOI: 10.2307 / 2404093
CrossRef Полный текст | Google Scholar
Норби, Р.Дж. И Ло Ю. (2004). Оценка реакции экосистемы на повышение содержания CO в атмосфере 2 и глобальное потепление в многофакторном мире. Новый Фитол . 162, 281–293. DOI: 10.1111 / j.1469-8137.2004.01047.x
CrossRef Полный текст | Google Scholar
Oono, Y., Seki, M., Satou, M., Iida, K., Akiyama, K., Sakurai, T., et al. (2006). Мониторинг профилей экспрессии генов Arabidopsis во время акклимации к холоду и деакклимации с использованием микрочипов ДНК. Funct. Интегр.Геномика 6, 212–234. DOI: 10.1007 / s10142-005-0014-z
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Пукацки П. М. и Каминска-Рожек Э. (2013). Реактивные виды, антиоксиданты и холодоустойчивость при дезакклимации Picea abies популяций. Acta Physiol. Завод . 35, 129–138. DOI: 10.1007 / s11738-012-1055-2
CrossRef Полный текст | Google Scholar
Репо, Т., Ханнинен, Х., Келломяки, С.(2006). Влияние длительного повышения температуры воздуха и СО на морозостойкость сосны обыкновенной. Среда растительных клеток . 19, 209–216. DOI: 10.1111 / j.1365-3040.1996.tb00242.x
CrossRef Полный текст | Google Scholar
Рииконен, Дж., Контунен-Соппела, С., Осипов, В., Тервахаута, А., Туомайнен, М., Оксанен, Э. и др. (2012). Метаболом хвои, морозостойкость и газообмен у проростков ели европейской, подвергшихся воздействию повышенной температуры и концентрации озона. Tree Physiol . 32, 1102–1112. DOI: 10.1093 / treephys / tps072
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Робинсон А. Р., Даув Р., Украинец Н. К., Каллис И. Ф., Уайт Р. и Мэнсфилд С. Д. (2009). Прогнозирование регенерационной способности соматических эмбриогенных культур хвойных с помощью метаболомики. Plant Biotechnol. J . 7, 952–963. DOI: 10.1111 / j.1467-7652.2009.00456.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Робинсон, А.Р., Украинец, Н. К., Канг, К.-Й., и Мэнсфилд, С. Д. (2007). Метаболитное профилирование пихты дугласовой ( Pseudotsuga menziesii ) в полевых испытаниях выявило сильную экологическую и слабую генетическую изменчивость. Новый Фитол . 174, 762–773. DOI: 10.1111 / j.1469-8137.2007.02046.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ресснер, У., Вагнер, К., Копка, Дж., Третуэй, Н., и Уиллмитцер, Л. (2000). Технический прогресс: одновременный анализ метаболитов в клубнях картофеля методом газовой хроматографии-масс-спектрометрии. Завод J . 23, 131–142. DOI: 10.1046 / j.1365-313x.2000.00774.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Rohloff, J., Kopka, J., Erban, A., Winge, P., Wilson, R.C., Bones, A.M, et al. (2012). Профилирование метаболитов выявило новые многоуровневые холодовые реакции у модели Fragaria vesca (лесная земляника). Фитохимия 77, 99–109. DOI: 10.1016 / j.phytochem.2012.01.024
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Ruelland, E., Vaultier, M.-N., Zachowski, A., and Hurry, V. (2009). Холодовая сигнализация и холодовая акклиматизация растений. Adv. Бот. Res . 49, 35–150. DOI: 10.1016 / S0065-2296 (08) 00602-2
CrossRef Полный текст | Google Scholar
Саид, А.И., Шаров, В., Уайт, Дж., Ли, Дж., Лян, В., Бхагабати, Н. и др. (2003). TM4: бесплатная система с открытым исходным кодом для управления и анализа данных микрочипов. BioTechniques 34, 374-378.
Pubmed Реферат | Pubmed Полный текст | Google Scholar
Сакаи, А.и Ларчер В. (1987). Морозостойкость растений: реакция и адаптация к морозному стрессу . Берлин; Гейдельберг: Springer Verlag. DOI: 10.1007 / 978-3-642-71745-1
CrossRef Полный текст | Google Scholar
Сакс, Х., Каннелл, М. Г. Р., Йонсен, О., Райан, М. Г., и Вурлитис, Г. (2001). Функционирование деревьев и лесов в ответ на глобальное потепление. Новый Фитол . 149, 369–399. DOI: 10.1046 / j.1469-8137.2001.00057.x
CrossRef Полный текст | Google Scholar
Ширяева, Л., Антти, Х., Шредер, В. П., Стримбек, Р., и Ширяев, А. С. (2012). Попарное многокомпонентное сравнение и OPLS-анализ фаз холодовой акклиматизации ели сибирской. Метаболомика 8, 123–130. DOI: 10.1007 / s11306-011-0304-5
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Sissener, N.H., Hemre, G.-I., Lall, S.P., Sagstad, A., Petersen, K., Williams, J., et al. (2011). Вызваны ли очевидные отрицательные эффекты кормления генетически модифицированной кукурузой MON810 атлантическому лососю, Salmo salar смешивающими факторами? Br.Дж. Нутр . 106, 42–56. DOI: 10.1017 / S0007114510005726
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Стримбек, Г. Р., Кьельсен, Т. Д., Шаберг, П. Г., и Мураками, П. Ф. (2008). Динамика низкотемпературной акклиматизации листвы хвойных пород умеренного и северного климата в условиях мягкого зимнего климата. Tree Physiol . 28, 1365–1374. DOI: 10.1093 / treephys / 28.9.1365
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Стримбек, Г.Р., Кьельсен, Т., Шаберг, П. Г., и Мураками, П. Ф. (2007). Холод в обычном саду: сравнительная низкотемпературная устойчивость хвойной листвы северного и умеренного климата. Деревья 21, 557–567. DOI: 10.1007 / s00468-007-0151-1
CrossRef Полный текст | Google Scholar
Стримбек, Г. Р., Шаберг, П. Г., Дехайес, Д. Х., Шейн, Дж. Б. и Хоули, Г. Дж. (1995). Среднезимнее закаливание ели горной красной во время естественной оттепели. Банка. J. Forest Res . 25, 2040–2044.DOI: 10.1139 / x95-221
CrossRef Полный текст | Google Scholar
Uleberg, E., Rohloff, J., Jaakola, L., Trôst, K., Junttila, O., Häggman, H., et al. (2012). Влияние температуры и фотопериода на урожай и химический состав северных и южных клонов черники ( Vaccinium myrtillus L.). J. Agric. Food Chem . 60, 10406–10414. DOI: 10.1021 / jf302924m
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Усадель, Б., Bläsing, O.E., Гибон, Y., Poree, F., Höhne, M., Günter, M., et al. (2008). Многоуровневый геномный анализ реакции транскриптов, активности ферментов и метаболитов в розетках Arabidopsis на постепенное снижение температуры в незамерзающем диапазоне. Среда растительных клеток . 31, 518–547. DOI: 10.1111 / j.1365-3040.2007.01763.x
Pubmed Реферат | Pubmed Полный текст | CrossRef Полный текст | Google Scholar
Вегис, А. (1964). Покой у высших растений.