строение и функции в процессе фотосинтеза — Природа Мира
Главная » Науки о природе
Время чтения 3 мин.Просмотры 25.3k.Обновлено
Фотосинтез происходит в эукариотических клеточных структурах, называемых хлоропластами. Хлоропласт — это тип органеллы растительных клеток, известный как зеленые пластиды. Пластиды помогают хранить и собирать необходимые вещества для производства энергии. Хлоропласт содержит зеленый пигмент, называемый хлорофиллом, который поглощает световую энергию для процесса фотосинтеза. Следовательно, название хлоропласт указывает на то, что эти органеллы представляют собой хлорофиллсодержащие пластиды.
Подобно митохондриям, хлоропласты имеют свою собственную ДНК, ответственны за производство энергии и воспроизводятся независимо от остальной части клетки посредством процесса деления, подобного бактериальному бинарному делению. Они также ответственны за производство аминокислот и липидных компонентов, необходимых для производства хлоропластов. Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласт: структура
Схема строения хлоропластХлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений. Охранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
- Мембрана — содержит внутренние и внешние липидные двухслойные оболочки, которые выступают в качестве защитных покрытий и сохраняют замкнутые структуры хлоропластов. Внутренняя мембрана отделяет строму от межмембранного пространства и регулирует прохождение молекул в/из хлоропласта.

- Межмембранное пространство — пространство между внешней и внутренней мембранами.
- Тилакоидная система — внутренняя система мембран, состоящая из сплющенных мешкообразных мембранных структур, называемых тилакоидами, которые служат местами преобразования энергии света в химическую энергию.
- Тилакоид с просветом (люменом) — отсек в каждом тилакоиде.
- Грана — плотные слоистые стопки тилакоидных мешков (10-20), которые служат местами преобразования энергии света в химическую энергию.
- Строма — плотная жидкость внутри хлоропласта, содержащая внутри оболочки, но вне тилакоидной мембраны. Здесь происходит конверсия углекислого газа в углеводы (сахара).
- Хлорофилл — зеленый фотосинтетический пигмент в хлоропласт-гране, поглощающий световую энергию.

Хлоропласт: фотосинтез
При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Гугломаг
Спрашивай! Не стесняйся!
Задать вопрос
Не все нашли? Используйте поиск по сайту
Search for:Хлоропласта структура
Хлоропласты у водорослей, как и у других растений, окружены оболочкой и состоят из основного вещества (стромы) и погруженных в него пластинчатых, или ламеллярных, структур, а также различного рода включений, отличающихся по размерам, форме и составу своего содержимого (рис. 6, 1и).[ …]
Хлоропласты обладают известной автономией в системе клетки. В них имеются собственные рибосомы и набор веществ, определяющих синтез ряда собственных белков хлоропласта. Имеются также ферменты, работа которых приводит к образованию липидов, входящих в состав ламелл, и хлорофилла. Как мы видели, хлоропласт располагает и автономной системой добывания энергии. Благодаря всему этому хлоропласты способны самостоятельно строить собственные структуры. Существует даже взгляд, что хлоропласты (как и митохондрии) произошли от каких-то низших организмов, поселившихся в растительной клетке и сперва вступивших с нею в симбиоз, а затем ставших ее составной частью, органоидом. [ …]
[ …]
| Структура хлоропластов. |
Изучение хлоропластов с помощью электронного микроскопа показало, что мембранная система имеет здесь дифференцированную, очень сложную, но четко упорядоченную структуру, которая различается у разных растений характером упаковки и степенью ее выраженности. Мембранная система погружена в строму (или матрикс) хлоропласта, в которой локализованы ферменты, связанные с восстановлением углекислоты и синтезом углеводов. Существенным обстоятельством является то, что в хлороплас-тах, кроме систем, непосредственно участвующих в осуществлении процесса фотосинтеза, имеется также собственная (отличная от ядерной) ДНК, рибосомы и другие компоненты белоксинтези-рующих систем. Для хлоропластов многих водорослей характерно также наличие специфического образования — пиреноида, функциональная роль которого остается неясной. Хлоропласты способны к делению и имеют чрезвычайно разнообразную форму у разных водорослей.[ … ]
]
| Схема структуры двойной ла-меллы хлоропласта |
Для развития хлоропластов из пропластид обычно необходим свет. Развитие происходит путем отшнурсмвывания от внутренней мембраны уплощенных пузырьков, образующих сплющенную, окруженную двойной мембраной пластинку тилакоидов, в которых в конечном итоге синтезируется хлорофилл. Пластиды, которые развиваются таким образом, ■ называются этиопластами.[ …]
В цитоплазме и хлоропластах клеток в зоне хлоротических местных некрозов, вызываемых ВТМ и огуречным вирусом 4, обнаружены группы аномальных окруженных мембранами пузырьков, причем в хлоропластах, как полагают, они образуются в результате выпячивания окружающих их мембран. Сферосомы клеток в зоне вокруг некротических местных поражений, образуемых ВТМ на листьях растений N. glutinosa, часто содержат одиночные включения с хорошо выраженной кристаллической структурой; в нормальных клетках подобные включения не обнаружены [855].[ …]
Па свету образуется хлорофилл. Короткие тилакоиды располагаются стопкой друг над другом и образуют тилакоиды гран. Пластиды достигают окончательного размера (рис. 30). Непосредственно ив про-пластид могут образовываться и бесцветные пластиды (лейкопласты— амплопласты) (рис. 31). Лейкопласты чаще всего локализованы в клетках запасающих тканей. Во многих случаях в лейкопластах ламеллы сохраняют связь с внутренней оболочкой. В строме лейкопластов располагаются крахмальные зерня, осмио-филыше глобулы, белковые включения. Эти глобулы располагаются сплошным слоем под оболочкой пластид.[ …]
Короткие тилакоиды располагаются стопкой друг над другом и образуют тилакоиды гран. Пластиды достигают окончательного размера (рис. 30). Непосредственно ив про-пластид могут образовываться и бесцветные пластиды (лейкопласты— амплопласты) (рис. 31). Лейкопласты чаще всего локализованы в клетках запасающих тканей. Во многих случаях в лейкопластах ламеллы сохраняют связь с внутренней оболочкой. В строме лейкопластов располагаются крахмальные зерня, осмио-филыше глобулы, белковые включения. Эти глобулы располагаются сплошным слоем под оболочкой пластид.[ …]
Для превращения пропластид в хлоропласта необходим свет. В темноте процессы синтеза и формирования мембранных структур прерываются.[ …]
Необходимо иметь в виду, что различные структуры, встречающиеся в незараженпой клетке, по своей морфологии могут напоминать вирусные включения. Так, в молодых хлоропластах корней фасоли найдены кристаллы запасного белка и связанные с ними пузырьки [1267]; кристаллические включения обнаружены в хлоропластах внешне здоровых листьев Масайатха [1360]. Маринос [1146] описал пластиды, содержащие вакуоли и мембрапы, в меристеме глазков клубней картофеля. Он назвал эти образования «ноли-функциопалыгами пластидами», но возможно, что некоторые из обнаруженных им ультраструктур возникли в результате «замаскированной» вирусной инфекции клубной.[ …]
Маринос [1146] описал пластиды, содержащие вакуоли и мембрапы, в меристеме глазков клубней картофеля. Он назвал эти образования «ноли-функциопалыгами пластидами», но возможно, что некоторые из обнаруженных им ультраструктур возникли в результате «замаскированной» вирусной инфекции клубной.[ …]
Резервирование (избыточность) субклеточных структур – очевидная вещь. Под структурной избыточностью понимают наличие в клетке функционально однозначных элементов в большем количестве, чем это необходимо для нормальной работы системы. Это уменьшает вероятность отказа системы в случае очень большой нагрузки на ее элементы. Примером является большое количество хлоропластов в клетке и их организация из множества равнозначных в функциональном отношении тилакоидов.[ …]
Хромопласты возникают либо из пропластид, либо из хлоропластов, либо из лейкопластов. Их внутренняя мембранная структура гораздо проще, чем у хлоропластов. Гран нет, строма содержит много желтого или оранжевого пигмента. Хромопласты содержатся в клетках лепестков, плодов, корнеплодов. [ …]
[ …]
На микрофотографиях показано (рис. 10 и 11), как изменяется структура тилакоидных систем листьев, подвергшихся газации. Тилакоиды хлоропластов контрольных растений в эксперименте 1 имеют форму продолговатых пузырьков. Тилакоиды у растений, подвергавшихся газации, в особенности при концентрации НС1 0,25 мг/м3 воздуха, характеризуются наличием регулярно повторяющихся электроноплотных участков, типичных для вполне развитых хлоропластов. Тилакоиды более отчетливо объединены в граны, но из-за неодинакового диаметра гран края их представляются заостренными. Вероятно, низкие концентрации НС1 ускоряют формирование гран.[ …]
При хроническом воздействии S02 наступают иные изменения структуры хлоропластов: матрикс стромы становится гранулярным и наблюдается аггломерация.[ …]
Показателями приема химических сигналов наружной мембраной хлоропласта или тилакоидной мембраной могут быть соответственно изменения ионной проницаемости или фотохимической активности, что позволяет характеризовать хлоропласт как биологическую структуру, подчиняющуюся единым закономерностям химической сигнализации. [ …]
[ …]
Болд и Солберг [67, 1640] с помощью фазово-контрастной микроскопии обнаружили структуры, названные ими «серыми пластинками», которые образуются на ранней стадии формирования кристаллов и которые, возможно, состоят из одиночного или двойного слоя ориентированных вирусных частиц. Вармке и Эдвардсон [1865] описали процесс роста кристаллов в клетках волосков табачного листа. На первых стадиях в цитоплазме инфицированных клеток вирусные частицы находились в свободном состоянии в форме небольших агрегатов параллельно расположенных палочек, соединяющихся торцам. Эти агрегаты затем увеличиваются в размерах и образуют структуры, аналогичные «серым пластинкам» Солберга и Болда. В процессе роста кристаллы не окружены какой-либо мембраной, и, поскольку они становятся многослойными, между их слоями оказываются иногда включенными часть эндоплазматической сети, митохондрии и даже хлоропласта.[ …]
Свечение обусловлено не фосфоресценцией, а особым строением пластинчатых структур протонемы, сидящих на прямостоячих ветвях, растущих в направлении света. Пластинка, вызывающая свечение, направлена против падающего света и слагается линзовидными клетками с выпуклой передней и воронковидной задней стенкой. Именно в заднем углу клетки размещены 4—6 хлоропластов. Луч света преломляется сферической передней стенкой, направляется на хлоропласты, пройдя насквозь хлоропласт, отражается задней стенкой; вновь преломленный в передней стенке, выходит параллельно входному лучу уже как зеленый свет. При изменении угла падения световых лучей изменяется и положение хлоропластов, собирающихся снова в фокусе преломленных лучей. Фокусированием световых лучей на хлоропластах достигается оптимальный для фотосинтеза эффект в условиях рассеянного света.[ …]
Пластинка, вызывающая свечение, направлена против падающего света и слагается линзовидными клетками с выпуклой передней и воронковидной задней стенкой. Именно в заднем углу клетки размещены 4—6 хлоропластов. Луч света преломляется сферической передней стенкой, направляется на хлоропласты, пройдя насквозь хлоропласт, отражается задней стенкой; вновь преломленный в передней стенке, выходит параллельно входному лучу уже как зеленый свет. При изменении угла падения световых лучей изменяется и положение хлоропластов, собирающихся снова в фокусе преломленных лучей. Фокусированием световых лучей на хлоропластах достигается оптимальный для фотосинтеза эффект в условиях рассеянного света.[ …]
Перекисная трансформация ненасыщенных жирных кислот, входящих в мембранные структуры, сопровождается нарушением полупроницаемости мембран, утратой способности к плазмолизу, высвобождением внутренних элементов клеточных органелл, нарушением осмотического равновесия, полной деструкцией гран хлоропластов. [ …]
[ …]
К этому подотделу относятся представители с одноклеточпым талломом монадной структуры (рис. 61). Продольная бороздка всегда тяпется вдоль брюшной стороны, в углублении на переднем конце клетки она переходит в треугольную глотку. Из глотки выходят два неодинаковых жгутика. Трихоцисты (рис. 61,1з, Зз) или слизистые тельца (рис. 61,4к), расположены в поверхностном слое цитоплазмы. Хлоропласты многочисленные, мелкие, дисковидные, светло-зеленые, между ними — капли масла.Пигментов меньше, чем в других группах, обнаружены только хлорофилл а, -каротин и несколько ксантофиллов. Есть и бесцветные формы. Вакуольный аппарат сложный. Ядро крупное. Запасное вещество — капли жира. Распространены преимущественно в сфагновых болотах, реже в озерах и других стоячих водоемах.[ …]
При прорастании зооспоры или зиготы сначала образуется сифонная булавовидная структура, ориентированная вертикально вверх. Прежде чем в ней начнется образование перегородок, она достигает заметных размеров, дифференцируясь на базальную (ризоидную) и апикальную части. Хлоропласт имеет сетчатое строение (рис. 236, 2), но, в отличие от остальных сифонокладовых, у кладофоры тенденция к соединению отдельных хлоропластов выражена в большей мере, поэтому часто хлоропласт выглядит как цельная, хотя и продырявленная пластинка, и увидеть, что он состоит из отдельных частей, можно лишь с помощью специальной цитологической техники.[ …]
Хлоропласт имеет сетчатое строение (рис. 236, 2), но, в отличие от остальных сифонокладовых, у кладофоры тенденция к соединению отдельных хлоропластов выражена в большей мере, поэтому часто хлоропласт выглядит как цельная, хотя и продырявленная пластинка, и увидеть, что он состоит из отдельных частей, можно лишь с помощью специальной цитологической техники.[ …]
Для растительной клетки характерно присутствие пластид. Важнейшие пластиды — это хлоропласты. Они осуществляют трансформацию световой энергии в химическую. Другой важнейший энергетический процесс (синтез АТФ за счет энергии окисления) происходит в митохондриях. Они представляют собой овальные или палочковидные структуры длиной 1…2 мкм. Система канальцев и цистерн (диктиосом), ограниченных однослойной мембраной, составляет аппарат Гольджи, основная функция которого — внутриклеточная секреция веществ, необходимых для построения клеточной оболочки и др. В округлых тельцах — лизосомах сконцентрированы гидролитические ферменты. С помощью сферосом идет синтез липидов.[ …]
С помощью сферосом идет синтез липидов.[ …]
Плазматическая мембрана, мембрана эндоплазматической сети, а также ядер, митохондрий и хлоропластов (см. ниже) представляют собой чрезвычайно сложные структуры, обладающие рядом важнейших биологических свойств. Многие мембраны содержат ферменты, транспортные системы, с помощью которых осуществляется перенос молекул питательных веществ и неорганических ионов внутрь клеток и внутри клеток, а также вывод из клеток продуктов жизнедеятельности. Мембранные структуры способны к самовосстановлению, если в них по каким-то причинам возникают повреждения.[ …]
У пораженных мучнистой росой растений наблюдается усиление транспирации и резкое ухудшение структуры хлоропластов в листьях. Недобор урожая может достигать 15 % и более.[ …]
Основная функция хлородластов — это процесс фотосинтеза. В 1955 г. Д. Ариоп показал, что в изолированных хлоропластах может быть осуществлен весь процесс фотосинтеза. Важво отметить, что хлоропласты имеются пе только в клетках листа. Последнее явление (хлорофиллопосность зародыша) привлекает внимание систематикой растений. Исследования покезалв. что структура хлоропластов, расположенных в других органах растения, так же как и состав пигментов, сходны с хлоронластами листа. Зто дает основания считать, что они способны к фотосинтезу. В том случае, если они подвергаются освещению, по-видимому, в них действительно происходит фотосинтез. Так, фотосинтез хлоропластов. расположенных в остях колоса, ыожет составлять около 30% от общего фотосинтеза растения. Позеленевшие па свету корни способны к фотосинтезу. В хлоропластах, находящихся, в кожуре плода до определенного этапа его развития, также может идти фотосинтез. Согласно предполвжению А. Л. Кур-санова, хлоропласты, расположенные вблизи проводящих путей, выделяя кислород, способствуют повышен иго интенсивности обмена веществ ситовидных трубок.[ …]
Последнее явление (хлорофиллопосность зародыша) привлекает внимание систематикой растений. Исследования покезалв. что структура хлоропластов, расположенных в других органах растения, так же как и состав пигментов, сходны с хлоронластами листа. Зто дает основания считать, что они способны к фотосинтезу. В том случае, если они подвергаются освещению, по-видимому, в них действительно происходит фотосинтез. Так, фотосинтез хлоропластов. расположенных в остях колоса, ыожет составлять около 30% от общего фотосинтеза растения. Позеленевшие па свету корни способны к фотосинтезу. В хлоропластах, находящихся, в кожуре плода до определенного этапа его развития, также может идти фотосинтез. Согласно предполвжению А. Л. Кур-санова, хлоропласты, расположенные вблизи проводящих путей, выделяя кислород, способствуют повышен иго интенсивности обмена веществ ситовидных трубок.[ …]
В клетках водорослей, так же как и в других зеленых растениях, но в отличие от грибных организмов, имеется хлоропласт-структура, обеспечивающая способность к фотосинтезу. [ …]
[ …]
По мере старения культуры водорослей и под действием токсиканта, в результате нарушения упорядоченности структуры хлоропластов и связанного с этим образования из хлорофилла различных продуктов деструкции, в клетках водорослей накапливаются некоторые количества феофитина. Продукты деградации хлорофилла в токсикологических опытах могут составлять значительную часть общего количества зеленых пигментов, что приводит к ошибкам в определении хлорофилла, поскольку они поглощают свет также в красной области спектра. Этот факт необходимо учитывать.[ …]
Достигнув определенного уровня, нарушения функции клеток могут отразиться на следующем, более высоком уровне организации. Изменения структуры ламеллярной системы хлоропластов, например, как видно на электрон-но-микроскопических фотографиях (рис. 10 и И), могут вызывать подавление восстановительных реакций в процессе ассимиляции СОг и задержку роста и развития, в особенности листьев; повреждение на уровне клетки может внешне проявляться в форме хлороза или некроза.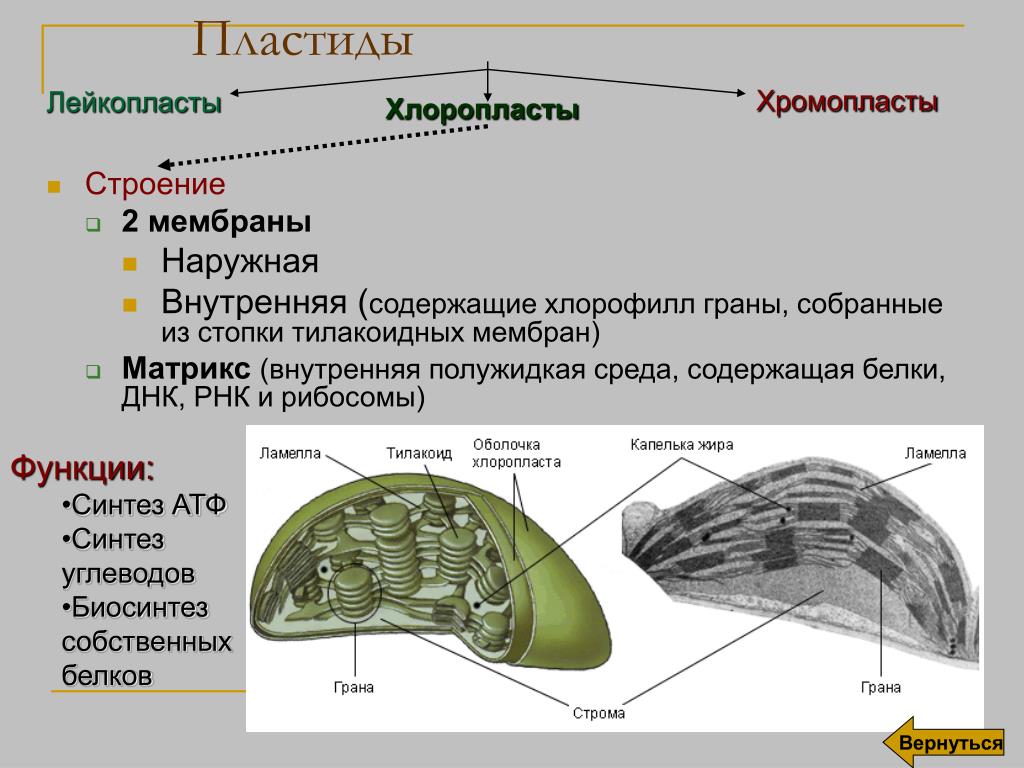 [ …]
[ …]
Т. Вейер предложил гранулярно-решетчатую модель, согласно которой внутренние пространства всех тнла-коидов соединены между собой (рис. 27). У хлоропластов большинства водорослей гран нет, а ламеллы собраны е группы (пачки) по 2—8 штук. Не во всех случаях и у Рис. 27. Схема гранулярно-сетча-высших растений хлоропласты Еме- той структуры хлоропласта.[ …]
В дальнейшем развитие клетки цошло не по линии совершенствования этого образования, а в направлении создания качественно иной, менее громоздкой структуры с аналогичными функциями, о чем свидетельствует отсутствие пиреноидов в хлоропластах растений, занимающих более высокую ступень развития по сравнению с водорослями.[ …]
Основой жизнедеятельности клетки являются обмен веществ и превращение энергии. Главным источником энергии на Земле является Солнце. Клетки растений специальными структурами в хлоропластах улавливают энергию Солнца, превращая ее в энергию химических связей молекул органических веществ и АТФ.[ . ..]
..]
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов — мостиков. Цитоплазматическая мембрана часто дает инвагинации — впячивания впутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной. Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы (рис. 2). Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур — аналоги митохондрий. Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки (табл. 30), которые по аналогии с гранулами хлоропластов растений называют стопками ти-лакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза. [ …]
[ …]
Возникновение у клетки способности делиться в двух и более плоскостях привело к образованию пластинчатой, мешковидной и трубчатой структуры весьма разнообразной формы (табл. 31, 2, 6, 7). Так же как у улотриксовых, слоевище ульвовых еще слабо дифференцировано. Заметно отличаются от остальных лишь крупные клетки в основании растения, снабженные ризоидными отростками, с помощью которых осуществляется прикрепление растений. Так же как у улотрикса, клетки ульвовых содержат единственный пристенный хлоропласт с одним или несколькими пиреноидами и одно ядро, расположенное по продольной оси клетки.[ …]
Однолетнее растение мятлик луговой (Роа annua) является одним из наиболее чувствительных к смогу, и его повсеместное распространение делает его весьма удобным индикатором смога. Поражения ограничены теми «летками, которые находятся в стадии максимального расширения. Они возникают на верхушке наиболее молодых листьев и постепенно переходят и на старые листья. Начальное и наиболее тяжелое поражение возникает в окружении подустьичных камер. За дезинтеграцией хлоропластов следует плазмолиз и, наконец, полная дегидратация пораженных клеток, что приводит к мумификации мезофильной ткани в пораженных участках.[ …]
За дезинтеграцией хлоропластов следует плазмолиз и, наконец, полная дегидратация пораженных клеток, что приводит к мумификации мезофильной ткани в пораженных участках.[ …]
Бактерии: прокариоты («доядерные») одноклеточные организмы. Их клетки не имеют отделенного от цитоплазмы ядра. Однако генетическая программа, как и у всех живых организмов, закодирована в виде последовательности нуклеотидов в ДНК и несет информацию о структуре белков. Бактериальные клетки не содержат таких органелл, как хлоропласты (специализированные для фотосинтеза) и митохондрии (специализированные для клеточного дыхания и синтеза АТФ). Эти биохимические процессы происходят у бактерий в цитоплазме.[ …]
С помощью люминесцентного микроскопа также изучают живые, нефиксированные клетки. Люминесценцией называется свечение объекта в результате поглощения световой энергии, вызываемое ультрафиолетовыми, а также синими и фиолетовыми лучами. Многие клеточные структуры обладают способностью к собственной (первичной) люминесценции. Так, хлорофилл, содержащийся в хлоропластах растительных клеток, обладает ярко-красной люминесценцией; довольно отчетливое свечение имеется у витаминов А и В, а также некоторых бактерий.[ …]
Так, хлорофилл, содержащийся в хлоропластах растительных клеток, обладает ярко-красной люминесценцией; довольно отчетливое свечение имеется у витаминов А и В, а также некоторых бактерий.[ …]
Недавно был предложен способ, который может помочь идентифицировать вирусные частицы в клетках, инфицированных мелкими сферическими вирусами 112181. Было найдено, что вирусоподобные частицы в растениях Chenopodium, обычно рассеянные в цитоплазме, образуют легко идентифицируемые кристаллические структуры, если перед фиксацией дать листьям слегка привянуть. Это наблюдение подкрепляет предположение о том, что образование кристаллических структур в инфицированных клетках зависит от концентрации вируса. Пользуясь такой методикой, мы не наблюдали больших количеств ВЖМТ-подобпых частиц внутри ядер и хлоропластов. Однако везикулярные структуры или внячивалия мембраны хлоронластов могут содержать ВЖМТ-подобные частицы. В клетках увядших листьев много таких частиц обнаруживалось также в пространствах между сгруппированными хлоропластами (эти пространства имеют примерно сферическую форму; фото 58) (Усияма и Мэтьюз, неопубликованные данные). Материал, заполняющий пространства между хлоропластами, включал большое количество меченого уридина [1036]. Эти эксперименты позволили нам условно сделать заключение, что местом сборки ВЖМТ в клетке является цитоплазма. Области цитоплазмы между сгруппированными хлоропластами, возможно, играют какую-то специфическую роль в качестве места синтеза и сборки ВЖМТ; однако этот вывод требует подтверждения.[ …]
Материал, заполняющий пространства между хлоропластами, включал большое количество меченого уридина [1036]. Эти эксперименты позволили нам условно сделать заключение, что местом сборки ВЖМТ в клетке является цитоплазма. Области цитоплазмы между сгруппированными хлоропластами, возможно, играют какую-то специфическую роль в качестве места синтеза и сборки ВЖМТ; однако этот вывод требует подтверждения.[ …]
Изучением диатомовых водорослей занимаются уже почти 150 лет, за этот период была предпринята не одна попытка их систематизации. Все системы диатомей можно разделить на три группы в зависимости от того, какие признаки были положены в их основу: 1) строение клеток и тип колоний; 2) положение, число и форма хлоропластов; 3) форма и детальная структура панциря. Системы первых двух групп оказались неудачными, так как форма колоний и хлоропластов в значительной мере зависит от условий среды и может часто меняться. Даже в пределах одного рода виды имеют различную форму хлоропластов и различное строение колоний в зависимости от того, где они обитают — в планктоне или бентосе. И наоборот, генетически не связанные между собой роды при наличии внешнего сходства по этим признакам включались в одну и ту же систематическую группу. Системы третьей группы базируются на более постоянных признаках — на строении панциря и деталях его структуры, позволяющих устанавливать родственные связи между таксонами. Кроме того, подобные системы имеют еще одно преимущество перед остальными: в них можно охватить не только современные, но и ископаемые формы, а это важно при создании действительно филогенетической системы диатомей.[ …]
И наоборот, генетически не связанные между собой роды при наличии внешнего сходства по этим признакам включались в одну и ту же систематическую группу. Системы третьей группы базируются на более постоянных признаках — на строении панциря и деталях его структуры, позволяющих устанавливать родственные связи между таксонами. Кроме того, подобные системы имеют еще одно преимущество перед остальными: в них можно охватить не только современные, но и ископаемые формы, а это важно при создании действительно филогенетической системы диатомей.[ …]
Если рассмотреть ранние стадии развития халимеды и удотеи (рис. 229, 7—11), то бросается в глаза родство всех сифоновых водорослей. Вначале зигота растет, оставаясь какое-то время одноядерной. Затем, на более поздней стадии, она претерпевает ряд цитологических и морфологических изменений. Одно крупное ядро делится, некоторые хлоропласты теряют свой крахмал и превращаются в лейкопласты, вся структура вытягивается в вертикальный сифон с первичным ризоидом в основании. Следующая стадия имеет разнонитчатое строение. И только на самых поздних стадиях развития происходит переплетение и смыкание вертикальных нитей.[ …]
Следующая стадия имеет разнонитчатое строение. И только на самых поздних стадиях развития происходит переплетение и смыкание вертикальных нитей.[ …]
В фазу колошения и созревания содержание марганца резко возрастает в колосе. Эти же авторы нашли, что марганец локализуется главным образом в незеленых цитоплазменных структурах и крупных фрагментах клетки. Так, по их данным, в крупных фрагментах клетки марганца содержится 325,1 мг на 100 г золы, или 16,3%; в хлоропластах 823,9, или 42,5%; в митохондриях 500, или 25,1%; в незеленых цитоплазменных структурах 321, или 16,1% марганца. Т.А.Парибок (1959) считает, что марганца в период вегетации больше в листьях, чем в стеблях. В период же созревания он мигрирует в колос, где содержание его возрастает до 38% от общего количества в растениях. Эти данные поволяют считать, что марганец имеет отношение в основном к функционирующим, деятельным органам растений.[ …]
Характер окрашивания этих трубочек наводит на мысль, что они содержат липиды или нуклеиновые кислоты [1216]. Принимая во внимание тот факт, что они появляются па очень рашшх стадиях инфекции, можно предположить, что они каким-то образом свя:шш с синтезом или сборкой вируса. Однако какими-либо определенными данными об их роли мы пока еще не располагаем. Подобные структуры наблюдали и другие авторы в клетках, инфицированных ВТМ. Было высказано предположение, что эти трубочки являются прямыми предшественниками палочек ВТМ; полагают, что палочки могли образовываться из них в результате процессов уплотнения и сжатия [1545]. В качестве главного возражения против такого вывода выдвигают в настоящее время различия в характере окрашивания этих трубочек и частиц ВТМ [1216], а также тот факт, что подобная стадия отсутствует при реконструкции ВТМ in vitro. Не удавалось наблюдать палочек ВТМ на топких срезах ядер или хлоропластов в инфицированных клетках [ 1213, 1545]. Однако хлоропласта в клетках больных растений могут образовывать нпячивашш или вакуолизироватьея, и в таких областях бывают видны палочки ВТМ.
Принимая во внимание тот факт, что они появляются па очень рашшх стадиях инфекции, можно предположить, что они каким-то образом свя:шш с синтезом или сборкой вируса. Однако какими-либо определенными данными об их роли мы пока еще не располагаем. Подобные структуры наблюдали и другие авторы в клетках, инфицированных ВТМ. Было высказано предположение, что эти трубочки являются прямыми предшественниками палочек ВТМ; полагают, что палочки могли образовываться из них в результате процессов уплотнения и сжатия [1545]. В качестве главного возражения против такого вывода выдвигают в настоящее время различия в характере окрашивания этих трубочек и частиц ВТМ [1216], а также тот факт, что подобная стадия отсутствует при реконструкции ВТМ in vitro. Не удавалось наблюдать палочек ВТМ на топких срезах ядер или хлоропластов в инфицированных клетках [ 1213, 1545]. Однако хлоропласта в клетках больных растений могут образовывать нпячивашш или вакуолизироватьея, и в таких областях бывают видны палочки ВТМ. Возможно, именно существованием таких впячиваний следует объяснить наличие небольших количеств ВТМ в препаратах изолированных хлоропластов. Локализация ВТМ в клетке может быть различной у разных штаммов вируса. Большая часть описанных выше работ выполнена на типичном штамме ВТМ. Однако для штамма UR, например, были получены четкие доказательства присутствия вируса в хлоропластах [1546]. Некоторые имеющиеся в литературе противоречия могут быть обусловлены различиями в поведении разных штаммов вируса.[ …]
Возможно, именно существованием таких впячиваний следует объяснить наличие небольших количеств ВТМ в препаратах изолированных хлоропластов. Локализация ВТМ в клетке может быть различной у разных штаммов вируса. Большая часть описанных выше работ выполнена на типичном штамме ВТМ. Однако для штамма UR, например, были получены четкие доказательства присутствия вируса в хлоропластах [1546]. Некоторые имеющиеся в литературе противоречия могут быть обусловлены различиями в поведении разных штаммов вируса.[ …]
Поверхностная мембрана — плазмалемма изолирует клетку от окружающей среды. Органеллы цитоплазмы имеют свои поверхностные мембраны. Вакуоль ограничена внутренней мембраной цитоплазмы — тоноплас-том. Мембраны составляют также внутреннюю структуру таких органелл, как хлоропласты и митохондрии, увеличивая поверхность, на которой протекают важнейшие биохимические и биофизические процессы.[ …]
На срезах листьев, находившихся в момент заражения в очень молодом возрасте, на которых развивалось типичное мозаичное заболевание, с помощью-светового микроскопа легко можно обнаружить ряд патологических отклонений в структуре хлоронластов. Островки ткани в мозаичных участках, имеющие различные оттенки зеленого, желтого и белого цветов, содержали, различные штаммы ВЖМТ, действующие на хлоропласта каждый своим особым и характерным образом (фото 37, цветная вклейка 1 и фото 56).[ …]
Островки ткани в мозаичных участках, имеющие различные оттенки зеленого, желтого и белого цветов, содержали, различные штаммы ВЖМТ, действующие на хлоропласта каждый своим особым и характерным образом (фото 37, цветная вклейка 1 и фото 56).[ …]
Однако в том виде, в каком они применялись, эти системы так и не дали возможности получить какие-либо определенные результаты. Объясняется это, вероятно, несколькими причинами. Во-первых, в растительных тканях, обычно присутствуют фенолы; они не только создают затруднения при выделении вирусов (гл. Во-вторых, в растительных тканях широко распространены нуклеазы, которые быстро инактивируют PIIK, если их но удалить или не подавить их активность. В-третьих, наличие у растений целлюлозных клеточных стенок означает, что для разрушения клеток требуются довольно жесткие методы. Между тем хлоропласта — хрупкие органеллы, и потому многие препараты, используемые в качестве бесклеточпых систем, загрязнены их фрагментами, которые очень трудно удалить. Наконец, в-четвертых, клетки взрослых, полностью развитых листьев имеют крупные вакуоли, содержимое которых может повреждать цитоплазматические структуры при разрушении клеток еще до того, как сыграют свою роль защитные вещества, присутствующие в экстрагирующей среде.[ …]
Наконец, в-четвертых, клетки взрослых, полностью развитых листьев имеют крупные вакуоли, содержимое которых может повреждать цитоплазматические структуры при разрушении клеток еще до того, как сыграют свою роль защитные вещества, присутствующие в экстрагирующей среде.[ …]
Хлоропласты и другие пластиды – Клетка
Хлоропласты, органеллы, ответственные за фотосинтез, во многом похожи на митохондрии. И хлоропласты, и митохондрии функционируют для выработки метаболической энергии, вырабатываемой эндосимбиозом, содержат свои собственные генетические системы и воспроизводятся путем деления. Однако хлоропласты крупнее и сложнее, чем митохондрии, и помимо образования АТФ они выполняют несколько важных задач. Самое главное, хлоропласты ответственны за фотосинтетическую конверсию CO 2 к углеводам. Кроме того, хлоропласты синтезируют аминокислоты, жирные кислоты и липидные компоненты собственных мембран. Восстановление нитрита (NO 2 – ) до аммиака (NH 3 ), важная стадия включения азота в органические соединения, также происходит в хлоропластах. Более того, хлоропласты являются лишь одним из нескольких типов родственных органелл (пластид), играющих разнообразные роли в растительных клетках.
Более того, хлоропласты являются лишь одним из нескольких типов родственных органелл (пластид), играющих разнообразные роли в растительных клетках.
Структура и функция хлоропластов
Хлоропласты растений представляют собой крупные органеллы (длиной от 5 до 10 мкм), которые, как и митохондрии, ограничены двойной мембраной, называемой оболочкой хлоропласта (). Помимо внутренней и внешней мембран оболочки хлоропласты имеют третью внутреннюю мембранную систему, называемую тилакоидной мембраной. Мембрана тилакоидов образует сеть уплощенных дисков, называемых тилакоидами, которые часто располагаются стопками, называемыми гранами. Из-за этой трехмембранной структуры внутренняя организация хлоропластов более сложна, чем у митохондрий. В частности, их три мембраны делят хлоропласты на три различных внутренних компартмента: (1) межмембранное пространство между двумя мембранами оболочки хлоропласта; (2) строма , которая лежит внутри оболочки, но снаружи тилакоидной мембраны; и (3) просвет тилакоидов.
Рисунок 10.13
Структура хлоропласта. В дополнение к внутренней и внешней мембранам оболочки хлоропласты содержат третью внутреннюю мембранную систему: тилакоидную мембрану. Эти мембраны делят хлоропласты на три внутренних отсека. (Электронная микрофотография (подробнее…)
Несмотря на большую сложность, мембраны хлоропластов имеют явное функциональное сходство с мембранами митохондрий — как и ожидалось, учитывая роль обеих органелл в хемиосмотическом образовании АТФ. оболочка хлоропластов, как и оболочка митохондрий, содержит порины и поэтому свободно проницаема для небольших молекул, в то время как внутренняя мембрана непроницаема для ионов и метаболитов, поэтому они могут проникать в хлоропласты только через специфические мембранные переносчики. и наружные мембраны оболочки хлоропласта сходны с внутренней и наружной мембранами митохондрий: в обоих случаях внутренняя мембрана ограничивает прохождение молекул между цитозолем и внутренней частью органеллы Строма хлоропласта также по функциям эквивалентна митохондриальной матрикс: Он содержит генетическую систему хлоропластов и множество метаболических ферментов, в том числе ответственных за критическое превращение CO 2 к углеводам при фотосинтезе.
Основное различие между хлоропластами и митохондриями с точки зрения как структуры, так и функции заключается в тилакоидной мембране. Эта мембрана имеет центральное значение в хлоропластах, где она выполняет роль внутренней митохондриальной мембраны в транспорте электронов и хемиосмотическом образовании АТФ (10). Внутренняя мембрана оболочки хлоропластов (не свернутая в кристы) не участвует в фотосинтезе. Вместо этого хлоропластная система переноса электронов расположена в мембране тилакоидов, и протоны перекачиваются через эту мембрану из стромы в просвет тилакоидов. Результирующий электрохимический градиент затем запускает синтез АТФ по мере того, как протоны возвращаются в строму. Таким образом, по своей роли в генерации метаболической энергии тилакоидная мембрана хлоропластов эквивалентна внутренней мембране митохондрий.
Рисунок 10.14
Хемиосмотическое образование АТФ в хлоропластах и митохондриях. В митохондриях транспорт электронов создает протонный градиент через внутреннюю мембрану, который затем используется для запуска синтеза АТФ в матриксе. В хлоропластах градиент протонов составляет (подробнее…)
В хлоропластах градиент протонов составляет (подробнее…)
Геном хлоропластов
Подобно митохондриям, хлоропласты содержат собственную генетическую систему, отражающую их эволюционное происхождение от фотосинтезирующих бактерий. Геномы хлоропластов сходны с геномами митохондрий тем, что они состоят из кольцевых молекул ДНК, присутствующих во множестве копий на органеллу. Однако геномы хлоропластов крупнее и сложнее, чем геномы митохондрий, размером от 120 до 160 т.п.н. и содержат примерно 120 генов.
Геномы хлоропластов нескольких растений были полностью секвенированы, что привело к идентификации многих генов, содержащихся в ДНК органелл. Эти хлоропластные гены кодируют как РНК, так и белки, участвующие в экспрессии генов, а также различные белки, участвующие в фотосинтезе. Как рибосомные, так и транспортные РНК, используемые для трансляции мРНК хлоропластов, кодируются геномом органеллы. К ним относятся четыре рРНК (23S, 16S, 5S и 4,5S) и 30 видов тРНК. В отличие от меньшего числа тРНК, кодируемых митохондриальным геномом, хлоропластных тРНК достаточно для трансляции всех кодонов мРНК в соответствии с универсальным генетическим кодом. Помимо этих РНК-компонентов системы трансляции, геном хлоропластов кодирует около 20 рибосомных белков, которые составляют примерно треть белков рибосом хлоропластов. Некоторые субъединицы РНК-полимеразы также кодируются хлоропластами, хотя дополнительные субъединицы РНК-полимеразы и другие факторы, необходимые для экспрессии генов хлоропластов, кодируются в ядре.
Помимо этих РНК-компонентов системы трансляции, геном хлоропластов кодирует около 20 рибосомных белков, которые составляют примерно треть белков рибосом хлоропластов. Некоторые субъединицы РНК-полимеразы также кодируются хлоропластами, хотя дополнительные субъединицы РНК-полимеразы и другие факторы, необходимые для экспрессии генов хлоропластов, кодируются в ядре.
Таблица 10.2
Гены, кодируемые хлоропластной ДНК.
Геном хлоропластов также кодирует около 30 белков, участвующих в фотосинтезе, в том числе компоненты фотосистем I и II, комплекса цитохрома bf и АТФ-синтазы. Кроме того, одна из субъединиц рибулозобисфосфаткарбоксилазы (rubisco) кодируется хлоропластной ДНК. Рубиско является критическим ферментом, который катализирует присоединение CO 2 к рибулозо-1,5-бисфосфату во время цикла Кальвина (см. ). Он не только является основным белковым компонентом стромы хлоропластов, но и считается самым распространенным белком на Земле, поэтому примечательно, что одна из его субъединиц кодируется хлоропластным геномом.
Импорт и сортировка белков хлоропластов
Хотя хлоропласты кодируют больше собственных белков, чем митохондрии, около 90% белков хлоропластов по-прежнему кодируются ядерными генами. Как и в случае с митохондриями, эти белки синтезируются на цитозольных рибосомах, а затем импортируются в хлоропласты в виде завершенных полипептидных цепей. Затем они должны быть отсортированы по соответствующему месту внутри хлоропластов — задача даже более сложная, чем сортировка белков в митохондриях, поскольку хлоропласты содержат три отдельные мембраны, которые делят их на три отдельных внутренних компартмента.
Импорт белка в хлоропласты обычно напоминает импорт митохондриального белка (). Белки предназначены для импорта в хлоропласты с помощью N-концевых последовательностей от 30 до 100 аминокислот, называемых транзитными пептидами
, которые направляют транслокацию белка через две мембраны оболочки хлоропласта и затем удаляются протеолитическим расщеплением. Транзитные пептиды распознаются транслокационным комплексом внешнего члена хлоропласта (Toc-комплекс), и белки транспортируются через этот комплекс через мембрану. Затем они переносятся в транслокационный комплекс внутренней мембраны (комплекс Tic) и транспортируются через внутреннюю мембрану в строму. Как и в митохондриях, молекулярные шапероны как на цитозольной, так и на стромальной сторонах оболочки необходимы для импорта белка, что требует энергии в виде АТФ. Однако, в отличие от предварительных последовательностей митохондриального импорта, транзитные пептиды не заряжены положительно, и транслокация полипептидных цепей в хлоропласты не требует наличия электрического потенциала через мембрану.
Затем они переносятся в транслокационный комплекс внутренней мембраны (комплекс Tic) и транспортируются через внутреннюю мембрану в строму. Как и в митохондриях, молекулярные шапероны как на цитозольной, так и на стромальной сторонах оболочки необходимы для импорта белка, что требует энергии в виде АТФ. Однако, в отличие от предварительных последовательностей митохондриального импорта, транзитные пептиды не заряжены положительно, и транслокация полипептидных цепей в хлоропласты не требует наличия электрического потенциала через мембрану.Рисунок 10.15
Белки, включенные в просвет тилакоидов, транспортируются к месту назначения в два этапа (). Сначала они импортируются в строму, как уже описано, а затем становятся мишенями для транслокации через тилакоидную мембрану с помощью второй гидрофобной сигнальной последовательности, которая экспонируется после расщепления транзитного пептида. Гидрофобная сигнальная последовательность направляет транслокацию полипептида через тилакоидную мембрану и, наконец, удаляется вторым протеолитическим расщеплением внутри просвета.
Гидрофобная сигнальная последовательность направляет транслокацию полипептида через тилакоидную мембрану и, наконец, удаляется вторым протеолитическим расщеплением внутри просвета.
Рисунок 10.16
Импорт белков в просвет тилакоидов. Белки импортируются в просвет тилакоидов в два этапа. Первым шагом является импорт в строму хлоропласта, как показано на рис. 10.15. Затем расщепление транзитного пептида обнажает второй гидрофобный (подробнее…)
Пути сортировки белков в другие четыре компартмента хлоропластов — внутреннюю и внешнюю мембраны, тилакоидную мембрану и межмембранное пространство — изучены менее четко. Как и в случае с митохондриями, белки, по-видимому, встроены непосредственно во внешнюю мембрану оболочки хлоропласта. Напротив, белки, предназначенные либо для тилакоидной мембраны, либо для внутренней мембраны оболочки хлоропласта, изначально становятся мишенями для импорта в строму с помощью N-концевых транзитных пептидов. После расщепления транзитных пептидов эти белки становятся мишенями для встраивания в соответствующую мембрану с помощью других последовательностей, которые еще недостаточно охарактеризованы. Наконец, не были идентифицированы ни последовательности, которые направляют белки в межмембранное пространство, ни пути, по которым они перемещаются к этому месту назначения.
Наконец, не были идентифицированы ни последовательности, которые направляют белки в межмембранное пространство, ни пути, по которым они перемещаются к этому месту назначения.
Другие пластиды
Хлоропласты являются лишь одним, хотя и наиболее заметным, членом большого семейства растительных органелл, называемых пластидами. Все пластиды содержат тот же геном, что и хлоропласты, но различаются как по структуре, так и по функциям. Хлоропласты специализируются на фотосинтезе и уникальны тем, что содержат внутреннюю тилакоидную мембранную систему. Другие пластиды, участвующие в различных аспектах метаболизма растительных клеток, ограничены двумя мембранами пластидной оболочки, но лишены как тилакоидных мембран, так и других компонентов фотосинтетического аппарата.
Различные типы пластид часто классифицируют по типам содержащихся в них пигментов. Хлоропласты названы так потому, что содержат хлорофилл. Хромопласты () лишены хлорофилла, но содержат каротиноиды; они ответственны за желтую, оранжевую и красную окраску некоторых цветов и фруктов, хотя их точная роль в клеточном метаболизме не ясна.
Рисунок 10.17
Электронные микрофотографии хромопластов и амилопластов. (A) Хромопласты содержат липидные капли, в которых хранятся каротиноиды. (B) Амилопласты содержат крупные гранулы крахмала. (A, Biophoto Associates/Photo Researchers, Inc.; B, Dr. Jeremy Burgess/Photo (подробнее…)
Все пластиды, включая хлоропласты, развиваются из пропластид , мелкие (от 0,5 до 1 мкм в диаметре) недифференцированные органеллы, присутствующие в быстро делящихся клетках корней и побегов растений. Затем пропластиды развиваются в различные типы зрелых пластид в соответствии с потребностями дифференцированных клеток. Кроме того, зрелые пластиды способны переходить из одного типа в другой. Хромопласты развиваются из хлоропластов, например, при созревании плодов (например, томатов).
Интересной особенностью пластид является то, что их развитие контролируется как сигналами окружающей среды, так и внутренними программами дифференцировки клеток. В фотосинтезирующих клетках листьев, например, пропластиды развиваются в хлоропласты (). В ходе этого процесса тилакоидная мембрана формируется везикулами, отпочковывающимися от внутренней мембраны пластидной оболочки, и синтезируются и собираются различные компоненты фотосинтетического аппарата. Однако хлоропласты развиваются только в присутствии света. Если растения содержатся в темноте, развитие пропластид в листьях останавливается на промежуточной стадии (называемой этиопластов ), в которых сформировался полукристаллический массив трубчатых внутренних мембран, но хлорофилл не синтезировался (). Если затем выращенные в темноте растения выставить на свет, этиопласты продолжают свое развитие до хлоропластов.
Рисунок 10.18
Развитие хлоропластов. Хлоропласты развиваются из пропластид в фотосинтезирующих клетках листьев. Пропластиды содержат только внутреннюю и внешнюю оболочечные мембраны; тилакоидная мембрана образуется путем отпочкования везикул от внутренней мембраны в процессе (подробнее…)
Рисунок 10.19
Электронная микрофотография этиопласта. (John N. A. Lott/Biological Photo Service.)
Хлоропласты – структура и функции
Присоединяйся сейчас
Если вы готовы сдать экзамены по биологии уровня A, станьте участником прямо сейчас, чтобы получить полный доступ ко всей нашей библиотеке материалов для повторения.
Присоединяйтесь к более чем 22 000 учащихся, сдавших экзамены благодаря нам!
Зарегистрируйтесь ниже, чтобы получить мгновенный доступ!
Присоединяйтесь →Или попробуйте пример…
Еще не готовы приобрести комплект для доработки? Без проблем. Если вы хотите ознакомиться с тем, что мы предлагаем, перед покупкой, у нас есть бесплатная подписка с образцами материалов для пересмотра.
Зарегистрируйтесь в качестве бесплатного члена ниже, и вы вернетесь на эту страницу, чтобы попробовать образцы материалов перед покупкой.
Скачать образцы →Quick Navigation
[скрыть]
Хлоропласты являются местом фотосинтеза в эукариотических клетках. Они присутствуют только в фотосинтезирующих клетках, таких как клетки растений и водорослей. В клетках животных и бактерий нет хлоропластов.
Структура хлоропластов- Хлоропласты высших растений обычно имеют двояковыпуклую или плосковыпуклую форму.
- Хлоропласты можно найти в клетках мезофилла листьев растений.
- У разных растений хлоропласты имеют различную форму: шаровидную, нитевидную, блюдцевидную, дисковидную или яйцевидную.
- Они везикулярные, с бесцветным центром.
- Некоторые хлоропласты имеют булавовидную форму, с тонкой средней зоной и заполненными хлорофиллом концами.
- У водорослей виден один огромный хлоропласт, который выглядит как сеть, спиральная полоса или звездчатая пластинка.
- Размер хлоропласта также варьируется от вида к виду и является постоянным для данного типа клеток.
- У высших растений средний размер хлоропластов составляет 4-6 мкм в диаметре и 1-3 мкм в толщину.

- Наружная мембрана – Это полупористая мембрана, проницаемая для небольших молекул и ионов, которые легко диффундируют. Наружная мембрана непроницаема для более крупных белков.
- Межмембранное пространство – Обычно это тонкое межмембранное пространство размером около 10-20 нанометров, которое находится между внешней и внутренней мембраной хлоропласта.
- Внутренняя мембрана – Внутренняя мембрана хлоропласта образует границу со стромой. Он регулирует прохождение материалов в хлоропласт и из него. Помимо регуляторной активности жирные кислоты, липиды и каротиноиды синтезируются во внутренней мембране хлоропластов.
- Строма- Строма представляет собой щелочную водную жидкость, богатую белком и присутствующую во внутренней мембране хлоропластов. Пространство за пределами тилакоидов называется стромой. ДНК хлоропластов, хлоропластные рибосомы и тилакоидная система, гранулы крахмала и многие белки обнаруживаются плавающими вокруг стромы.
- Тилакоидная система — Тилакоидная система подвешена в строме. Система тилакоидов представляет собой набор мембранных мешочков, называемых тилакоидами. Хлорофилл содержится в тилакоидах и служит средством для протекания процесса световых реакций фотосинтеза. Тилакоиды расположены стопками, известными как граны. Каждая гранула содержит около 10-20 тилакоидов.


- Поглощение световой энергии и преобразование ее в биологическую энергию.
- Производство NAPDh3 и выделение кислорода в процессе фотолиза воды.
- Производство АТФ фотофосфорилированием. НАДФ3 и АТФ являются ассимиляционными силами фотосинтеза. Перенос СО2, полученного из воздуха, на 5-углеродный сахар в потоке во время темновой реакции.
- Расщепление 6-атомного соединения на две молекулы фосфоглицериновой кислоты за счет использования ассимиляционных сил.
- Преобразование PGA в различные сахара и хранение в виде стрейча. Хлоропласт очень важен, так как он является местом приготовления пищи для всех зеленых растений. Все гетеротрофы также зависят от пластов для этой пищи.
- У растений все клетки участвуют в иммунном ответе растений, так как у них отсутствуют специализированные иммунные клетки.
- Хлоропласты с ядром и клеточной мембраной и ЭР являются ключевыми органеллами защиты от патогенов.
- Наиболее важной функцией хлоропластов является производство пищи в процессе фотосинтеза.
- Пища готовится в виде сахаров. В процессе фотосинтеза сахар и кислород производятся с использованием энергии света, воды и углекислого газа.
- Световые реакции происходят на мембранах тилакоидов.
- Хлоропласты, как и митохондрии, используют потенциальную энергию ионов Н+ или градиент ионов водорода для выработки энергии в форме АТФ.
- В строме хлоропластов происходят темновые реакции, также известные как цикл Кальвина.
- Образование молекул НАДФ3 и кислорода в результате фотолиза воды.
- Путем использования ассимиляционных сил 6-атом углерода расщепляется на две молекулы фосфоглицериновой кислоты.

Часто задаваемые вопросы
Что такое хлоропласты?
Хлоропласты представляют собой связанные с двойной мембраной органеллы, встречающиеся только в растительных клетках. Они участвуют в процессе фотосинтеза.