цвет, фото, текстура, свойства, применения
Ель относится к семейству “сосновые” и насчитывает почти 40 видов вечнозелёных деревьев, высота которых достигает 30 метров.
Еловые леса находятся на третьем месте по занимаемой площади среди хвойных пород деревьев, уступает ель лишь лиственницам и сосновым.
Чистые еловые леса имеют плотную, густую и тёмную структуру.Древесина ели мягкая, лёгкая, не обладает высокой прочностью, используется в качестве строительного материала (доски, брусья), для небольших изделий, а также для переработки в древесную массу.
Основной особенностью структуры ели можно назвать то, что она не обладает чётко выраженным ядром. Древесина имеет однородный бело-желтоватый цвет, редко с бледным розовым оттенком. Годичные кольца можно увидеть в любом разрезе.
Использование ели в качестве пиломатериалов происходит вместе с пихтой, потому что обе породы имеют схожие качества. Из еловой древесины изготавливают доски, шпон,фанеру,склеенный брус, ДСП и ДВП.
В строительстведомовом ель используется при изготовлении крыш, деревянной обшивки, перил, лестниц, каркасов стен и потолков, полов, окон, дверей и ворот. Доска из ели применяется также при изготовлении подоконников, настилов полов, выполнении обшивочных и декоративных работ. Она имеет приятный запахи желто-розовый цвет с кремовым оттенком.
В результате наличия высокого содержания различных смол обрезная доска из ели не гниет, влагостойка и выдерживает высокую температуру. Использовать ее можно где угодно, кроме возведения бань, потому что при высоких температурах будет происходить выделение смолы в воздух.
Области применения ели
Древесина ели легче сосновой и хорошо колется. В прошлом она широко использовалась для получения колотых кровельных материалов: гонта и дранки. По гибкости Древесины ель превосходит другие хвойные породы, кроме можжевельника. Поэтому ее древесина используется для производства гнутых изделий (дуги, сани, охотничьи лыжи и др.).
Большая длина трахеид делает еловую древесину ценным сырьем для производства бумаги. Кора ели тонкая и в сыром состоянии эластичная. Она используется на кровли временных построек. В коре много дубителей, но по качеству они хуже дубителей лиственничной и дубовой коры.
Кора ели тонкая и в сыром состоянии эластичная. Она используется на кровли временных построек. В коре много дубителей, но по качеству они хуже дубителей лиственничной и дубовой коры.
Долинные ельники предотвращают водную эрозию и играют кольматирующую роль – фильтруют талые и дождевые воды от взвешенных в них твердых частиц, чем предохраняют водоемы от заиления.
| Научная классификация | Физические свойства | ||
| Домен: | Эукариоты | Средняя плотность: | 430 кг/м³ |
| Царство: | Растения | Пределы плотности: | |
| Отдел: | Хвойные | Продольная усадка: | 0,3 % |
| Класс: | Хвойные (Pinopsida Burnett, 1835) | Радиальная усадка: | 3,6 % |
| Порядок: | Сосновые | Тангенциальная усадка: | 7,8 % |
| Семейство: | Сосновые | Радиальное набухание: | 0,19 % |
| Род: | Ель | Тангенциальное набухание: | 0,36-0,39 % |
| Международное научное название | Прочность на сгиб: | 68 Н/мм² | |
|
Picea A. |
Прочность на сжатие: | 40 Н/мм² | |
| Типовой вид | Предел прочности: | 80 Н/мм² | |
| Picea rubra A.Dietr. = Picea abies (L.) H.Karst. — Ель обыкновенная | Топливные свойства | ||
| 20,2 МДж/кг | |||
 Dietr., 1824
Dietr., 1824Виды ели
Род Ель, по данным Королевских ботанических садов Кью, включает 37 первичных видов и 4 гибридогенных:
- Picea abies (L.) H.Karst. — Ель обыкновенная, или Ель европейская
- Picea alcoquiana (H.J.Veitch ex Lindl.) Carrière — Ель Алькокка, или Ель двуцветная
- Picea asperata Mast. — Ель шероховатая
- Picea aurantiaca Mast.
- Picea austropanlanica Silba
- Picea brachytyla (Franch.) E.Pritz. — Ель китайская
- Picea breweriana S.
 Watson — Ель Бревера
Watson — Ель Бревера - Picea chihuahuana Martínez
- Picea crassifolia Kom.
- Picea engelmannii Parry ex Engelm. — Ель Энгельмана
- Picea farreri C.N.Page & Rushforth
- Picea glauca (Moench) Voss — Ель белая, или Ель сизая
- Picea glehnii (F.Schmidt) Mast. — Ель Глена
- Picea jezoensis (Siebold & Zucc.) Carrière — Ель аянская
- Picea koraiensis Nakai — Ель корейская
- Picea koyamae Shiras.
- Picea likiangensis (Franch.) E.Pritz. — Ель ликиангская
- Picea linzhiensis (W.C.Cheng & L.K.Fu) Rushforth
- Picea mariana (Mill.) Britton, Sterns & Poggenb. — Ель чёрная
- Picea martinezii T.F.Patt.
- Picea maximowiczii Regel ex Mast. — Ель Максимовича
- Picea meyeri Rehder & E.H.Wilson — Ель Мейера
- Picea morrisonicola Hayata — Ель мориссонская
- Picea neoveitchii Mast. — Ель новая Вича
- Picea obovata Ledeb. — Ель сибирская
- Picea omorika (Pančić) Purk. — Ель сербская
- Picea orientalis (L.) Peterm. — Ель восточная
- Picea polita (Siebold & Zucc.) Carrière — Ель изящная, или Ель японская (или Picea torano (Siebold ex K. Koch) Koehne, о легитимности первого названия до сих пор ведётся дискуссия)
- Picea pungens Engelm. — Ель колючая, или Ель голубая
- Picea purpurea Mast. — Ель пурпурная
- Picea retroflexa Mast. — Ель отогнутая
- Picea rubens Sarg. — Ель красная
- Picea schrenkiana Fisch. & C.A.Mey.— Ель Шренка
- Picea schrenkiana subsp. tianschanica (Rupr.) Bykov — Ель тянь-шанская
- Picea sitchensis (Bong.) Carrière — Ель ситхинская
- Picea smithiana (Wall.) Boiss.
- Picea spinulosa (Griff.) A.Henry — Ель шиповатая, или Ель восточно-гималайская
- Picea wilsonii Mast. — Ель Вильсона
 Watson — Ель Бревера
Watson — Ель Бревера — Ель сибирская
— Ель сибирская- Гибридогенные виды
- Picea ×albertiana S. Br.
- Picea ×fennica (Regel) Kom. — Ель финская (Picea abies × Picea obovata)
- Picea ×lutzii Little
- Picea ×notha Rehder (Picea glehnii × Picea jezoensis)
 Br.
Br.Полезные таблицы
Различия в характеристиках древесины и основные сферы применения сосны и ели
| Основные характеристики древесины |
Сосна |
Ель |
|---|---|---|
| Цвет | Красноватый или желтоватый (охровый) оттенок | Белая |
| Стабильность цвета | Со временем оттенок становится более интенсивным | Долго сохраняется естественный светлый цвет |
| Структура | Ярко выражена | Слабо выражена, высокая равномерность расположения волокон. |
| Плотность при влажности 12% | 480 кг/куб.м. | 430 кг/куб. м. м. |
| Смолистость | Высокая | Ниже средней |
| Сучки | Небольшое количество, крупные и очень крупные, овальной формы, располагаются группами. | Большое количество, мелкие, круглые |
| Стойкость к загниванию | Высокая | Средняя |
| Стойкость к посинению (свежесрубленная древесина) | Низкая | Высокая |
| Прочность | Высокая | Несколько меньше, чем у сосны |
| Свойства, имеющие значение для механической обработки | Мягкая, легко обрабатывается, не растрескивается при высыхании | Мягче сосны, но сучки затрудняют обработку, мало подвержена короблению |
| Сфера использования |
|
|
Значения коэффициента влагопроводности (Dх1010 м2/с) для ели
|
Спелая древесина: |
200С |
600С |
800С |
|
тангенц. |
2,65 |
9,05 |
17,7 |
|
радиальное напр. |
2,78 |
9,2 |
20,0 |
|
Заболонь: |
200С |
600С |
800С |
|
тангенц.напр. |
3,16 |
12,1 |
19,0 |
|
радиальное напр. |
3,26 |
13,7 |
19,6 |
 напр.
напр.ЕЛЬ • Большая российская энциклопедия
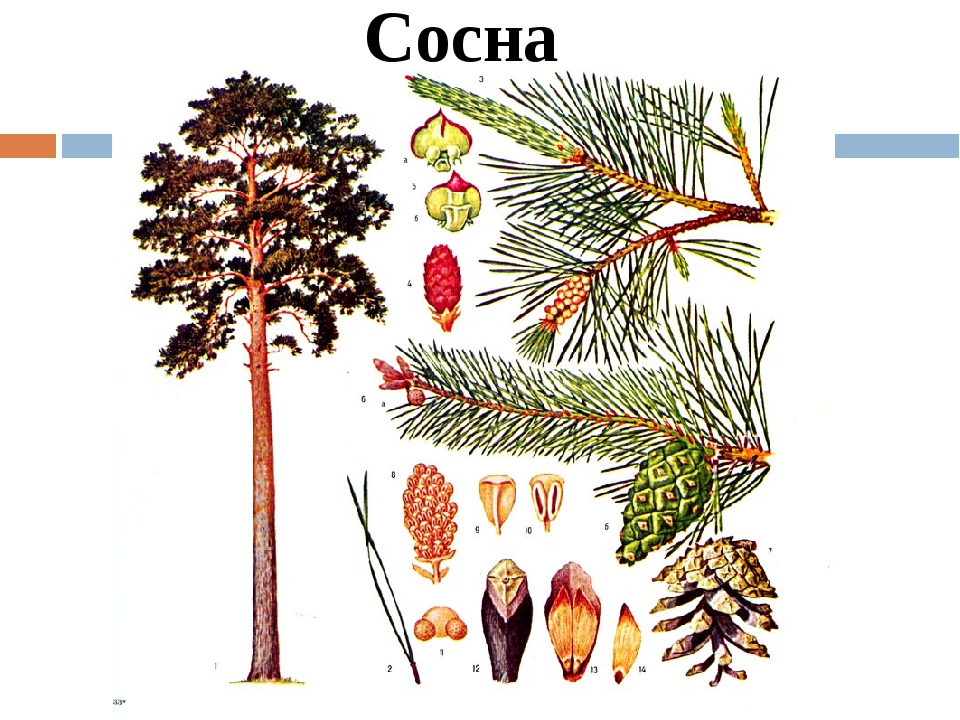
Ель обыкновенная: 1 – общий вид; 2 – ветвь с мужскими шишечками; 3 – зрелая женская шишка; 4 – семя.
ЕЛЬ (Picea), род голосеменных хвойных растений сем. сосновых. Достоверные ископаемые остатки Е. обнаружены в олигоценовых отложениях. Как правило, это однодомные вечнозелёные деревья с высокими прямыми стволами и почти мутовчато расположенными ветвями, образующими густую конич. крону, однако на верхней границе леса в горах и у сев. границы леса растения нередко принимают стланиковую (стелющуюся) форму. В среднем высота Е. составляет 30–40 м, но Е. аянская (P. ajanensis) и сев.-амер. Е. ситхинская (P. sitchensis) достигают 50–70 м и более (самые крупные в мире Е. ситхинские могут достигать 96,7 м в высоту и 5,72 м в диаметре). Листья игольчатые (хвоя), четырёхгранные, спирально расположенные на побегах, не опадают в течение 7–8 лет и более; их длина до 2,5 см, с одним проводящим пучком и двумя смоляными каналами. Пазушные микростробилы (мужские шишки) собраны на побегах предыдущего года и несут многочисл. микроспорофиллы с парными пыльцевыми мешками. Пыльцевые зёрна с двумя возд. мешками. Деревянистые мегастробилы (женские шишки) у разных видов от 2 до 15 см длиной, висячие, целиком опадающие после высыпания семян. Семена 4–5 мм длиной, с плёнчатым асимметричным крылом, созревают обычно на второй год.
Пазушные микростробилы (мужские шишки) собраны на побегах предыдущего года и несут многочисл. микроспорофиллы с парными пыльцевыми мешками. Пыльцевые зёрна с двумя возд. мешками. Деревянистые мегастробилы (женские шишки) у разных видов от 2 до 15 см длиной, висячие, целиком опадающие после высыпания семян. Семена 4–5 мм длиной, с плёнчатым асимметричным крылом, созревают обычно на второй год.
Ок. 40 видов, распространённых в холодных, умеренных, а также горных субтропич. областях Евразии и Сев. Америки. Наибольшее видовое разнообразие – в Центр. и Зап. (горном) Китае и в Сев. Америке. Во флоре России 7 видов. В таёжной зоне Е. – важнейшие лесообразующие породы, слагающие темнохвойные леса равнин и горно-таёжные леса среднего горного пояса. В Европ. части России осн. лесообразующей породой темнохвойных (ельников) и смешанных лесов является Е. обыкновенная, или европейская (P. abies, или P. excelsa), выс. до 30–40 м (иногда до 52 м) и 1–1,5 м в диаметре. Кора серая, отслаивающаяся у старых деревьев тонкими чешуйками. Корневая система поверхностная, поэтому при сильном ветре деревья нередко выворачиваются с корнями. Начинает давать семена с 15, а в насаждениях – с 25–30 лет. Зрелые женские шишки коричнево-каштановые, блестящие, 10–15 см длиной и 3–4 см шириной; семенные чешуи ромбовидные, вверху зубчатые. Обыкновенная Е. отличается высокой теневыносливостью, морозоустойчива, выносит избыточное увлажнение, но страдает от засух, поздних весенних и ранних осенних заморозков, копоти, заводских газов, сухости воздуха. Живёт 250–300, редко до 450 лет. Растёт на свежих глинистых, суглинистых и супесчаных почвах. На северо-востоке Европ. части, по всей Сибири (исключая тундровые районы) и далее до побережья Охотского м.
обыкновенная, или европейская (P. abies, или P. excelsa), выс. до 30–40 м (иногда до 52 м) и 1–1,5 м в диаметре. Кора серая, отслаивающаяся у старых деревьев тонкими чешуйками. Корневая система поверхностная, поэтому при сильном ветре деревья нередко выворачиваются с корнями. Начинает давать семена с 15, а в насаждениях – с 25–30 лет. Зрелые женские шишки коричнево-каштановые, блестящие, 10–15 см длиной и 3–4 см шириной; семенные чешуи ромбовидные, вверху зубчатые. Обыкновенная Е. отличается высокой теневыносливостью, морозоустойчива, выносит избыточное увлажнение, но страдает от засух, поздних весенних и ранних осенних заморозков, копоти, заводских газов, сухости воздуха. Живёт 250–300, редко до 450 лет. Растёт на свежих глинистых, суглинистых и супесчаных почвах. На северо-востоке Европ. части, по всей Сибири (исключая тундровые районы) и далее до побережья Охотского м. и ср. Амура широко распространена Е. сибирская (P. obovata), с более мелкими шишками, цельнокрайными и закруглёнными наверху семенными чешуями. На севере и северо-востоке европ. тайги обитает Е. финская (P. fennica), сформировавшаяся путём гибридизации двух вышеупомянутых видов. В среднегорном поясе Зап. Кавказа произрастает Е. восточная (P. orientalis), достигающая 57 м в высоту и 4 м в диаметре; на Дальнем Востоке – Е. корейская (P. koraiensis), Е. Глена (P. glehnii) и важнейшая из доминант темнохвойных лесов Дальнего Востока – Е. аянская.
и ср. Амура широко распространена Е. сибирская (P. obovata), с более мелкими шишками, цельнокрайными и закруглёнными наверху семенными чешуями. На севере и северо-востоке европ. тайги обитает Е. финская (P. fennica), сформировавшаяся путём гибридизации двух вышеупомянутых видов. В среднегорном поясе Зап. Кавказа произрастает Е. восточная (P. orientalis), достигающая 57 м в высоту и 4 м в диаметре; на Дальнем Востоке – Е. корейская (P. koraiensis), Е. Глена (P. glehnii) и важнейшая из доминант темнохвойных лесов Дальнего Востока – Е. аянская.
Е. играет важнейшую роль в лесном хозяйстве России. Её лёгкая и мягкая древесина применяется в строительстве, столярном и мебельном произ-ве, целлюлозно-бумажной пром-сти, при изготовлении муз. инструментов. Из неё добывают смолу, дёготь, скипидар, канифоль, древесный уксус. Кора Е., содержащая таннины, используется при дублении кож. Хвоя, богатая каротином, витамином С и эфирным маслом, перерабатывается на хвойно-витаминную муку (добавка в корм скоту и домашней птице). Настой и отвары хвои применяют в нар. медицине (обладают противоцинготным, мочегонным и противомикробным действием). Еловая смола в смеси с воском и сливочным маслом – прекрасное средство от ссадин и ран. Е. декоративны, их разводят в парках, скверах, на улицах, высаживают в лесных полосах и вдоль железных и шоссейных дорог. Помимо отеч. видов, в ландшафтном строительстве популярны сев.-амер. виды и формы Е. с серебристой и голубоватой хвоей – Е. Энгельманна (P. engelmannii), Е. сизая (P. glauca) и Е. колючая (P. pungens).
Хвоя, богатая каротином, витамином С и эфирным маслом, перерабатывается на хвойно-витаминную муку (добавка в корм скоту и домашней птице). Настой и отвары хвои применяют в нар. медицине (обладают противоцинготным, мочегонным и противомикробным действием). Еловая смола в смеси с воском и сливочным маслом – прекрасное средство от ссадин и ран. Е. декоративны, их разводят в парках, скверах, на улицах, высаживают в лесных полосах и вдоль железных и шоссейных дорог. Помимо отеч. видов, в ландшафтном строительстве популярны сев.-амер. виды и формы Е. с серебристой и голубоватой хвоей – Е. Энгельманна (P. engelmannii), Е. сизая (P. glauca) и Е. колючая (P. pungens).
Теперь каждым Рождественским британским людям посылают огромную ель из Норвегии, которая стоит в Квадрате Trafalgar, в центре Лондона, сияя вниз на всех людях, которые собираются в Сочельник. | |
| Люди украшают свои дома и, как правило, ставят ель, которая стоит в углу комнаты. | |
| Он повернулся спиной и уселся у огня на поваленную ветром ель. | |
| Это было поваленное дерево прямо посреди дороги в заповедную часть Беловежской пущи. Через эту ель с трудом перебрался директорский джип. | |
| Столетнюю ель скрутило как обычную ветку. | |
| В середине 1990-х годов сосна обыкновенная, бук, дуб каменный, ель обыкновенная, европейский и скальный дуб восстановились после своей первоначальной дефолиации. | |
| Шагах в двадцати от саней стояла громадная засохшая ель. | |
| Индейка, ель, эгног бабушки Эстер, и всё такое. Мы будем рады, если ты придёшь. | |
| Слова: бор, буря, ведьма, ель, Еж, мрак, мосток, медведь, метель | |
Хозяин кафе раздобыл четырехметровую ель, которую водрузили около стойки. Роза, признанный авторитет по части уюта и задушевной атмосферы, взялась украсить дерево. | |
| Ель в лесу, под елью белка, Белка песенки поет | |
| Добравшись до цели, он огляделся вокруг, высматривая, где больше хвороста, чтобы свалить ель в ту сторону. | |
| Скажу тебе вот что: если Лемон не получит Рождественскую ель, самолюбие будет последним, о чём стоит беспокоиться. | |
| Они забрали ель и носки. | |
| Я имею ввиду, что норвежская ель достаточно быстрорастущее хвойное, я знаю, ты пытаешься отвлечь меня, пока я поднимаюсь, но я собираюсь сделать это в любом случае, так что.. | |
| Каждый раз, когда человек выдергивал хворост из кустов, ель слегка сотрясалась – едва заметно для него, но достаточно сильно, чтобы вызвать катастрофу. | |
| Глаза Шеннон зажглись, как рождественская ель. | |
| Ель растет перед дворцом. | |
| Трудно украшать ель без ели, м? | |
Помните большую ель, которая подпирала нашу кладовку со стороны реки? – начал Элия. | |
| Но Ель должно быть хвастается этим почему-то. | |
| Встречу мне, хрипя, поднималась мать, протягивая сухие руки без мяса на них, длинная, тонкая, точно ель с обломанными ветвями. | |
| Разве я не упомянула, что ты поступила в ель благодаря этим деньгам? | |
| Ель Морадо был в 30 милях через Рио-Гранде. | |
| Любовник Юлии Вольмар, Малек-Ад ель и де Линар, | |
| Даже элитные гильдии вовсю ищут эту самую ель. | |
| Или, может, Вы хотите серебристую ель? | |
| Когда я подошла к вашему окошку, я уже подозревала… Вы только представьте себе: сегодня ночью, когда Ад ель возвращалась домой, я услышала, кроме ее шагов, еще какие-то мужские. | |
| О ты внесла Драма школу в список ЗА Ель? | |
| Все что хотят эти коралевы – золотой билет в Ель. | |
Были посажены многие другие виды хвойных деревьев, включая Дугласову ель и лиственницу, но они были менее терпимы к местным условиям, чем сосны. | |
| Лучше всего растут ясень, вяз, обыкновенный бук, дуб и серебристая ель. | |
| Старый Тикко-это 9.550-летняя норвежская ель, расположенная на горе Фулуфьеллет провинции Даларна в Швеции. | |
| Кроме того, использовались ясень, вяз, сосна, ель и лиственница. | |
| Хотя он используется для лонжеронов в наше время, пока нет никаких доказательств, что викинги использовали ель для мачт. | |
| Самые громкие деревянные вершины, такие как ель Ситка, легкие и жесткие, сохраняя при этом необходимую прочность. | |
| Черная ель является значительным деревом в пределах водораздела реки Теслин. | |
| Некоторые виды 8 миллионов деревьев Виннипега включают вяз, ясень, клен, дуб, тополь, липа, ива, береза, ель, сосна, кедр, а также некоторые фруктовые деревья и кустарники. | |
Ель, например, очень распространена, но большие куски с ровным зерном составляют небольшую долю от общего запаса и могут быть дорогими. | |
| 56 процентов территории Латвии занимают леса, в основном сосна Шотландская, береза и ель норвежская. | |
| В более влажных районах можно встретить клен, ель, сосну, ель и дуб. | |
| Ель обыкновенно росла среди других засухоустойчивых степных трав, таких как бутелуа хирсута и Хилария белангери. | |
| Наиболее распространенными породами деревьев являются сосна, ель и береза. | |
| В более высоких зонах преобладают хвойные леса-сосна Шотландская, Сосна Боснийская, сосна македонская, пихта болгарская, пихта Серебряная, ель норвежская и др. | |
| Вечная мерзлота мешает деревьям развивать глубокие корневые системы; например, черная ель, которая адаптировалась к вечномерзлым почвам, не имеет значительного стержневого корня. | |
| В горах чаще встречаются ель, пихта и сосна. | |
Picea glauca, белая ель, – это вид ели, произрастающий в северных умеренных и бореальных лесах Северной Америки. | |
| Белая ель может жить несколько сотен лет. | |
| Смола блистера, как правило, не хватает, но Porsild ель обыкновенная Глаука ВАР. | |
| Белая ель имеет трансконтинентальный ареал в Северной Америке. | |
| Белая ель – самый северный вид деревьев в Северной Америке, достигающий чуть севернее 69°северной широты в дельте реки Маккензи. | |
| Белая ель обычно встречается в регионах, где вегетационный период превышает 60 дней в год. | |
| Как экзотика, белая ель широко распространена, но встречается редко. | |
| Белая ель-это кульминационное пологое дерево в бореальных лесах Канады и Аляски. | |
| Белая ель чаще всего произрастает в почвах порядков Альфизолов и Инцептизолов. | |
| Белая ель будет расти в зонах выращивания USDA 3-7, но не приспособлена к жаре и влажности и будет плохо работать в жарком климате. | |
Белая ель становится менее приспособленной к почвенным условиям с увеличением суровости климата. | |
| Белая ель чрезвычайно вынослива к низким температурам, при условии, что растение находится в состоянии зимнего покоя. | |
| Белая ель подвержена сильному повреждению от весенних заморозков. | |
| На большей части своего ареала белая ель встречается чаще в ассоциации с деревьями других видов, чем в чистых древостоях. | |
| Белая ель имеет большое экономическое значение в Канаде из-за ее древесины, собранной для производства бумаги и строительства. | |
| Белая ель-это дерево провинции Манитоба и дерево штата Южная Дакота. | |
| Изоргапонтин можно найти в еловых породах, таких как белая ель. | |
| Норвежская ель, сербская ель, Шотландская сосна и турецкая пихта не являются коренными жителями Соединенных Штатов. | |
| Восточная белая сосна родом из восточных штатов США, а Колорадская голубая ель-из Колорадо и Юты. | |
Белая ель произрастает на севере центральной части США и Канады. | |
| Самый распространенный вид, который выращивают как свежесрезанные деревья Фрейзер ель, сосна обыкновенная, псевдотсуга, пихта бальзамическая. | |
| Райты построили самолет в 1903 году, используя в качестве строительного материала гигантскую ель. | |
| Искусственная елка-это искусственная сосна или ель, изготовленная специально для использования в качестве рождественской елки. | |
| Пихта была измерена на уровне 15%, красное дерево-15%, дуб-14%, ель, 3.40%, 5.80%, 7.70%, 9.95%, 17.0% и еще 16%. | |
| Другие результаты | |
Ель обыкновенная | Лесная кладовая
Здравствуйте, дорогой читатель!
Ель обыкновенная – самое распространенное хвойное дерево в западном секторе лесной зоны Евразии. Проще говоря, это обычная наша елка, прекрасно всем знакомая. Но и в знакомом, привычном, будничном можно находить новое и неизвестное.
Ель обыкновенная, или европейская
Обыкновенную ель называют еще елью европейской. Хотя в Западной и Центральной Европе дерево растет только в горах. Наиболее же обычна эта ель в Северной Европе, Беларуси, на севере Украины. И, конечно, на севере Европейской России, где она формирует значительные лесные массивы.
Хотя в Западной и Центральной Европе дерево растет только в горах. Наиболее же обычна эта ель в Северной Европе, Беларуси, на севере Украины. И, конечно, на севере Европейской России, где она формирует значительные лесные массивы.
На востоке, ближе к Уралу, и на самом севере лесной зоны ель обыкновенная сменяется близким видом – елью сибирской. Вид близкий, но все же другой – с более короткой и колючей хвоей, более мелкими шишками, меньшей высотой. И способностью выживать в более суровом климате.
Вид другой, но все же близкий. Ели обыкновенная и сибирская скрещиваются, образуя жизнеспособные гибриды. Говорят даже об особом переходном виде – ели финской.
Если внимательно рассмотреть шишки елей обыкновенной и сибирской, можно заметить различия, считающиеся видовыми признаками. Край чешуи у ели сибирской закругленный и гладкий, а у обыкновенной — с небольшими зубчиками, зазубринками.
Ель относят к семейству сосновых. В самом деле, несмотря на очевидные различия, у этих деревьев немало общего. Кроме зеленой хвои, сохраняющейся несколько лет, ель обыкновенную объединяет с сосной двудомность – на одном дереве созревают и мужские, и женские шишки. Строение и происхождение шишек, строение пыльцы и семян, процессы, протекающие при опылении и оплодотворении, также сходны.
Кроме зеленой хвои, сохраняющейся несколько лет, ель обыкновенную объединяет с сосной двудомность – на одном дереве созревают и мужские, и женские шишки. Строение и происхождение шишек, строение пыльцы и семян, процессы, протекающие при опылении и оплодотворении, также сходны.
Различий много. В отличие от сосны, ели способны вырастать высокими и стройными деревьями независимо от того, растут ли они в густом лесу или же на открытом месте. Дело в том, что нарастает ель обыкновенная преимущественно своей верхушечной почкой. Именно она дает самые длинные побеги – от 30 до 50 см ежегодно.
Причем растет ель своей верхушкой всю жизнь. Правда, при условии – если верхушечная почка не повреждена. Или несущий эту почку побег почему-либо не удален. В этом случае функции верхушечной берет на себя одна из боковых почек. Но дерево уже никогда не вырастет высоким и стройным.
Верхушку ели всегда венчает «корона» из почек: одна верхушечная и несколько боковых. Весной они дают побеги. И образуется мутовка. Так же как у сосны обыкновенной. И возраст молодой ели также легко определить, сосчитав число этих мутовок и прибавив 5 – 7 лет. В течение первых лет жизни мутовки на дереве не образуются.
И образуется мутовка. Так же как у сосны обыкновенной. И возраст молодой ели также легко определить, сосчитав число этих мутовок и прибавив 5 – 7 лет. В течение первых лет жизни мутовки на дереве не образуются.
Боковые ветви тоже ежегодно нарастают, но значительно меньше верхушки. Причем и на боковой ветви ели каждый год подрастают боковые побеги – уже относительно самой этой ветви. Это тоже мутовки, только не полные – веточки отходят не во все стороны, а близко к одной плоскости. Образуется еловая ветвь, которую мы обычно называем еловой лапой.
Побеги у ели обыкновенной, в отличие от сосны, только одного типа – удлиненные. Напомню, что у сосны обыкновенной, кроме ежегодно нарастающих удлиненных побегов, есть еще укороченные, длиной всего пару миллиметров. На них растет пара сосновых хвоинок. Вместе с хвоей побеги эти и опадают через 2 – 3 года, или чуть больше.
Еловая хвоя растет прямо на удлиненном побеге. Хвоинки, значительно короче сосновых, усеивают весь побег, располагаясь по спирали. Сидит хвоинка на листовой подушечке. Когда она опадет, на коре остается листовой след.
Сидит хвоинка на листовой подушечке. Когда она опадет, на коре остается листовой след.
Еловая хвоя уплощенно-четырехгранная, с колючей верхушкой. Длина хвои 1 – 2 см. Держится на дереве она дольше сосновой. В естественных условиях срок жизни хвоинки составляет до 10 – 12 лет. Правда, у деревьев, растущих в условиях повышенного загрязнения воздуха, хвоясменяется значительно раньше.
Ель обыкновенная, как и другие представители этого рода, неплохо переносит затенение. Поэтому даже в густом еловом лесу крона дерева остается весьма развитой. Засыхают от недостатка света лишь самые нижние ветви. Крона ели, растущей на открытом месте, обычно пирамидальная. Ветви растут на стволе почти до самой земли.
Развитая крона хорошо обеспечивает дерево питательными веществами. Ведь чем больше на дереве листьев (хвоинок), тем больше производится сахаров при фотосинтезе. Но такая крона может доставить дереву и серьезные проблемы.
Зимой у нас выпадает много снега. Даже лишенные листьев березы под его тяжестью частенько гнутся, а то и ломаются. Ели обыкновенной обильные снегопады больших неприятностей не доставляют. Тонкие, но прочные и гибкие ветви под тяжестью снега тоже сгибаются. И сбрасывают его!
Ели обыкновенной обильные снегопады больших неприятностей не доставляют. Тонкие, но прочные и гибкие ветви под тяжестью снега тоже сгибаются. И сбрасывают его!
Но вот сильные ветры при большой парусности кроны часто выворачивают дерево целиком. Способствуют этому и особенности корневой системы ели. Только лет до пятнадцати у дерева растет стержневой корень. А затем активно разрастаются корни боковые, залегающие в верхнем слое почвы. Удержать высокое дерево при сильном ветре такие корни не могут. И рушатся лесные великаны.
Живет ель европейская до 250 – 300 лет. Вот только встретить в лесу такие деревья вряд ли удастся. Разве где-то в заповеднике. Большинство елей вырубается, не доживая и до столетнего юбилея.
Никогда не рубленный ельник оставляет впечатления незабываемые! Приходилось мне побывать в таком лесу много лет назад. Это на северо-западе Вологодской области, почти на границе с Карелией, в верховьях реки Андомы. Ассоциации … сказочные. Кажется, вот-вот из-за соседнего дерева выглянет Баба-Яга. Или Леший.
Или Леший.
На десятки метров вверх уходят мощные колонны елей. Их диаметр у комля – более метра. Ветви увешаны бородами лишайника уснеи. Тихо в таком лесу и сумрачно. Почва, валежник, включая целые стволы упавших от старости или ветра огромных елей – все покрыто толстым слоем зеленых мхов. Из кустарничков растет разве что черника, да и то не везде.
Там, где посветлее – возле лесного ручья, к примеру – появляются и некоторые травы. Сверкают белые звездочки седмичника европейского. А в местах близкого залегания грунтовых вод зеленые мхи сменяются болотным мхом сфагнумом.
На свежих пнях на просеке под лесовозную дорогу, что достигла тогда уже этих мест, можно подсчитать годичные кольца, что не преминули сделать ботаники нашей экспедиции. Насчитывали 250 – 300 колец.
По итогам экспедиции, в которой я тогда работал, был создан Верхнеандомский государственный заказник. Массив коренных ельников взяли под охрану. Что там теперь – сказать не могу …
Ель обыкновенная гораздо требовательнее сосны к почвенным условиям.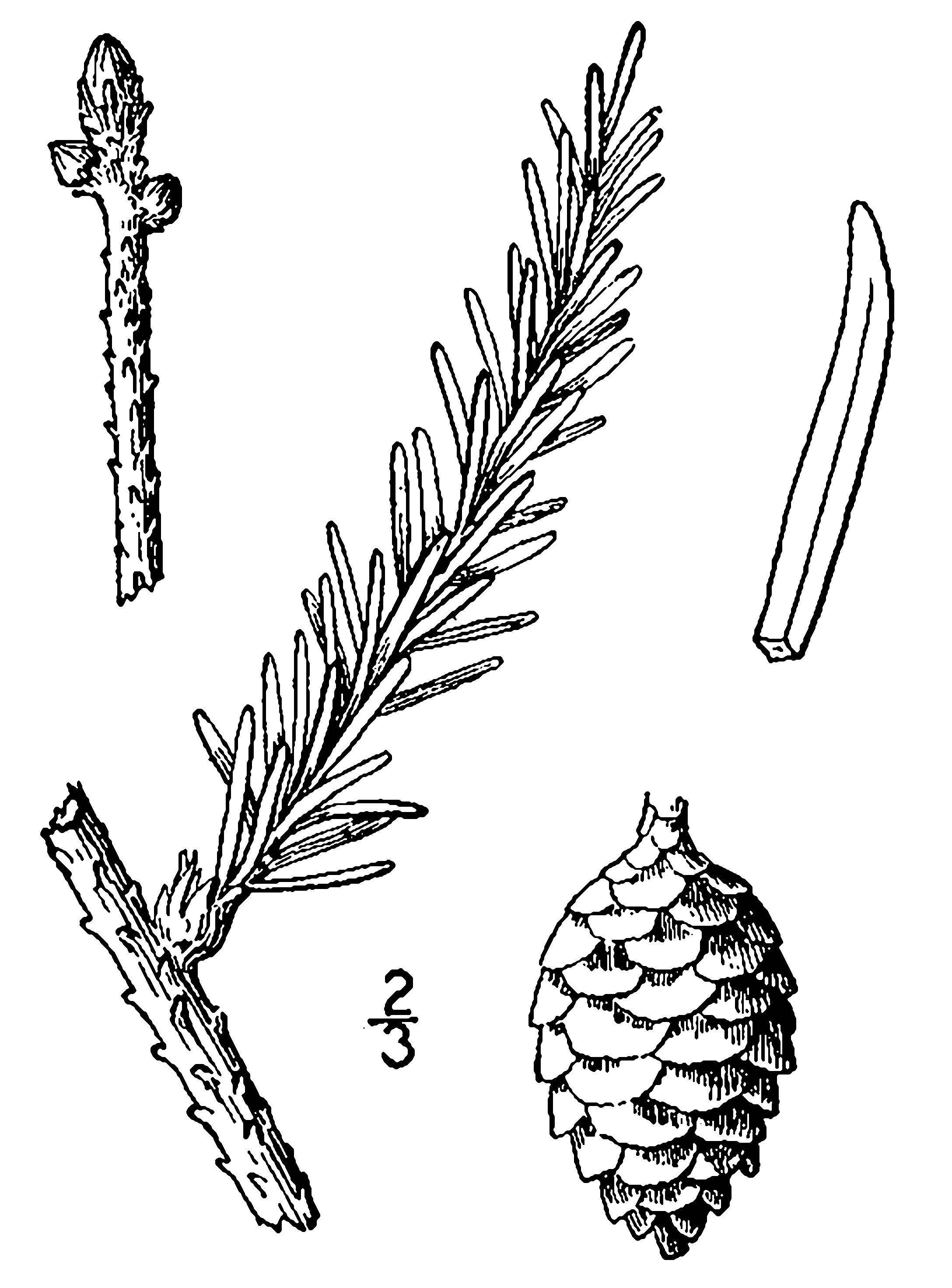 Она не будет расти ни на сухих песках, ни на верховом болоте. А еще она плохо переносит засухи. Потому уже на юге лесной зоны встречается реже.
Она не будет расти ни на сухих песках, ни на верховом болоте. А еще она плохо переносит засухи. Потому уже на юге лесной зоны встречается реже.
Ель обыкновенная весной
Зиму деревья проводят в состоянии своеобразной «спячки», когда жизненные процессы замедляются. Не исключение и деревья хвойные. Устьица на хвое плотно закрыты – нужно экономить воду. Корни не могут в достатке обеспечить ею дерево, воду в холодной почве корешки практически не всасывают.
Впрочем, при температурах выше – 5 градусов в хвое все же начинается фотосинтез. Но такие температуры для наших зим не характерны.
Но вот приходит весна, и все начинает быстро меняться. Еще на рубеже сезонов, во время, поэтично названное М.М.Пришвиным «весной света», в сухие солнечные дни вскрываются еловые шишки, высыпая семена, разносимые ветром. В мае, с приходом тепла, сначала набухают, а потом распускаются почки, давая начало новым вегетативным побегам.
Рассмотрите еловые лапы в это время. На концах веточек набухли крупные почки, прикрываемые бледно-желтыми колпачками почвенных чешуй. Кое-где чешуйки эти уже раздвинулись, а то и опали. Из-под них появляется на свет кисточка светло-зеленых иголочек. Это молодой побег.
Кое-где чешуйки эти уже раздвинулись, а то и опали. Из-под них появляется на свет кисточка светло-зеленых иголочек. Это молодой побег.
Молодые иголочки отличаются от старых не только по цвету. Они мягкие и совершенно не колючие. Если же «кисточку» сорвать и разжевать, ощущается кисленький вкус. И никакого смолистого привкуса и аромата.
Молодые побеги быстро растут. В мае – начале июня они еще отличаются от старых цветом своей хвои. Но с приходом настоящего лета рост побегов прекращается, хвоя твердеет и приобретает обычные свойства.
Почти одновременно с вегетативными распускаются и генеративные почки. Из них появляются видоизмененные побеги ели обыкновенной – ее женские и мужские шишки. Ель «зацветает». Происходит это почти одновременно с цветением черемухи.
Конечно, биологи поправляют – хвойные не цветут, у них нет цветка. Но все же сходство большое, особенно если учесть, что шишки в это время выглядят весьма зрелищно.
Несколько подробнее о «цветении» ели есть отдельная статья здесь.
Рассмотреть молодые шишки ели обычно довольно сложно, так как располагаются они в верхней части кроны. Разве что повезет … На верхушках прошлогодних побегов появились небольшие желтенькие или красноватые мужские шишки (или мужские колоски). В мешочках под чешуями созревает огромное количество пыльцы.
Пыльцевые зерна ели обыкновенной, подобно сосновым, имеют воздушные мешки, благодаря чему их удельный вес мал. Пыльца далеко уносится ветром, покрывает листья деревьев, траву. Если пройдет дождь, желтая пыльца хорошо заметна на лужах.
Женские еловые шишки, величиной вначале с наперсток, развиваются на верхушках побегов, которым уже два года. Эти «наперсточки» растут на ветке вертикально. Цвет молодых шишек малиновый или ярко-красный. Подрастая, они все больше напоминают некие «новогодние украшения», красивые свечи.
После опыления шишка закрывает свои чешуи, закупоривает проходы между ними смолой. Меняет она и свое положение на ветке – теперь шишка не «торчит», а свисает. Цвет она тоже меняет – становится зеленой или зеленовато-бурой.
Цвет она тоже меняет – становится зеленой или зеленовато-бурой.
Любопытные особенности жизни ели обыкновенной
Ели не нужно, как сосне, двух лет на развитие семян. Шишки ее созревают в этот же год, осенью. К этому времени еловые шишки достигли своего максимального размера в 10 – 15 см. Они стали коричневыми и блестящими. Семена в шишках созрели, но они не вскрываются. Дерево будет ждать новой «весны света».
Зимой еловые семена служат отличной пищей многим лесным обитателям. Белки, дятлы, клесты обрывают шишки, доставая из них семена. Шишки, брошенные или упавшие на землю, служат добычей наземным грызунам – лесным мышам.
Правда, хороший урожай еловых шишек бывает не ежегодно. Такие годы повторяются лишь через 5 – 7 лет. Прошедший год был именно таким. Сейчас на елках висит огромное количество шишек. Но они уже пусты, без семян.
В конце зимы шишки вскрываются, отправляя семена в полет. Как и семя сосны, еловое семечко снабжено крылышком. Падая в воздухе, семена кружатся подобно маленьким «вертолетикам», что замедляет падение и позволяет улететь дальше от материнского дерева.
Из семени вырастет крохотный проросток, мало напоминающий будущее дерево. Далеко не каждое семечко прорастет, особенно в густом ельнике. Ведь на поверхности почвы здесь лежит толстый слой лесной подстилки, состоящий в основном из опавшей хвои. Проростку очень трудно пробиться к почве своим слабым корешком. Лесные мхи тоже могут оказаться непреодолимым препятствием. Потому семян нужно много.
Но ель обыкновенная способна прорастать в несколько неожиданных местах. В лесу обычно много валежника, и слой мхов, тем более хвои, покрыть его не могут. Еловое семечко, попадая на такую полусгнившую валежину или на пень, прорастает.
Лесоводы ради обеспечения естественного возобновления ели используют прием, называемый сдиранием подстилки. Удаляя в некоторых местах слой лесной подстилки и зеленых мхов, они дают дополнительный шанс семенам ели обыкновенной на прорастание.
Жизнь молодого деревца под пологом елового леса порой весьма нелегка. Не хватает самого важного – света. Ведь в густом ельнике почти не бывает подлеска – деревьев и кустарников нижних ярусов. Им здесь просто не выжить из-за недостатка света. Даже мхи порой не растут – лишь толстый слой опавшей хвои покрывает землю.
Ведь в густом ельнике почти не бывает подлеска – деревьев и кустарников нижних ярусов. Им здесь просто не выжить из-за недостатка света. Даже мхи порой не растут – лишь толстый слой опавшей хвои покрывает землю.
Еловый подрост тоже угнетен, растет плохо. Но все же держится! Годичные кольца у такой елочки, не достигающей роста человека, порой можно рассмотреть только под лупой. Но результат подсчета может ошеломить – «деревце» бывает и пятидесятилетним, и даже старше!
Его ровесники, растущие в более благоприятных условиях, уже стали мощными высокими деревьями. А елочка все ждет своего часа. И часто дожидается. Рухнул от старости или под напором ветра один из лесных великанов. В образовавшуюся прогалину хлынул свет. И еловый подрост начинает стремительно расти, за считанные годы достигая верхнего яруса леса.
То же самое произойдет, если древостой (взрослые деревья верхнего яруса) вырубается человеком. При одном условии – если при вырубке не пострадает подрост. Именно поэтому лесоводы требуют от лесозаготовителей сохранения подроста. Ведь еловый лес, выросший естественным образом, значительно качественнее выращенного руками человека.
Ведь еловый лес, выросший естественным образом, значительно качественнее выращенного руками человека.
На открытом месте молодым елочкам хорошо. И одновременно … плохо. Света здесь хватает. Зато и открыта елочка всем капризам погоды, включая сильные весенние заморозки.
Если заморозок случится, когда молодые побеги тронутся в рост, то они могут серьезно пострадать. Нежная молодая хвоя повреждается даже легким морозцем. И стоят в начале лета такие елочки, будто огнем опаленные.
А вот под пологом березы, ольхи серой или осины елочкам действительно комфортно. И света хватает, и от весеннего морозца прикрывают. Да и почва в местах произрастания лиственных деревьев обычно плодородная.
Лет до пятнадцати и здесь ель обыкновенная все же растет медленно. После же начинает набирать рост ускоренными темпами, догоняя, а потом и обгоняя лиственные деревья. Когда же над большей частью березовой рощи сомкнутся еловые кроны, березняк обречен.
Старые березы со временем отомрут. А подрост березы в густой тени ели расти не может. Происходит смена формаций – березняки и осинники заменяются ельниками.
А подрост березы в густой тени ели расти не может. Происходит смена формаций – березняки и осинники заменяются ельниками.
Ель обыкновенная – одно из самых полезных для человека деревьев в наших лесах. Но об этом – в другой раз, в отдельной статье.
Если Вы все еще не подписаны на новости блога «Лесная кладовая», рекомендую сделать это прямо сейчас. Впереди еще много интересного и полезного!
С уважением, Александр Силиванов
Розовые шишки ели • Яна Домбровская • Научная картинка дня на «Элементах» • Ботаника
На фото — молодая женская шишка обыкновенной ели, она розовая и ее чешуи призывно раскрыты в ожидании пыльцы. По-научному шишки голосеменных растений называются стробилы. Женские шишки — это мегастробилы: у ели они вырастают до 15–16 см. Мужские шишки — микростробилы: они маленькие, всего пару сантиметров длиной.
Обыкновенная ель живет до нескольких сотен лет: самому старому известному дереву 468 лет, а некоторым корням шведской ели Старый Тикко — 9500 лет. Размножаться же она начинает в возрасте 20–60 лет: в мае на концах ее ветвей вырастают те самые розовые мегастробилы. Предположительно, такой относительно темный цвет обеспечивает лучшее поглощение тепла в северных широтах. До опыления эти шишки задорно ориентированы вверх. Чешуи их открыты и готовы принимать пыльцу.
Размножаться же она начинает в возрасте 20–60 лет: в мае на концах ее ветвей вырастают те самые розовые мегастробилы. Предположительно, такой относительно темный цвет обеспечивает лучшее поглощение тепла в северных широтах. До опыления эти шишки задорно ориентированы вверх. Чешуи их открыты и готовы принимать пыльцу.
Одновременно с этим на боковых побегах созревают микростробилы. Они представляют собой ось с отходящими вбок редуцированными спороносными листьями — микроспорофиллами. На них развиваются микроспорангии, в которых формируются микроспоры. Из микроспоры развивается мужской гаметофит (гаплоидная, то есть с одинарным набором хромосом, стадия жизненного цикла растений) — пыльцевое зерно, или пылинка. Гаметофит несет две мужские гаметы — спермии. В ясную погоду из микростробилов высыпаются мириады созревших пыльцевых зерен. Пылинки имеют два воздушных мешка, которые придают им парусность и позволяют перемещаться на большие расстояния с помощью ветра. Они могут утонуть в ближайшей луже, а могут достичь своей цели — мегастробила.
Мегастробил состоит из оси и плотно прилегающих к ней чешуй. Чешуи двух типов: мелкие кроющие (наружные) и более крупные семенные (внутренние), они растут одна над другой. Соответственно, наружная чешуя играет защитную роль, а на внутренней формируется два семязачатка. В отличие от мужских шишек, чешуи женских — это не спорофиллы, а укороченные видоизмененные побеги (мегаспорофиллы есть только у саговников).
Семязачаток состоит из оболочки с отверстием в верхней части (это отверстие называется микропиле, или пыльцевход), расположенной под ним пыльцевой камеры и мегаспорангия — нуцеллуса. В нуцеллусе образуется четыре мегаспоры, но созревает только одна, остальные отмирают. Из мегаспоры развиваются женский гаметофит эндосперм и запасающая ткань (в клетках эндосперма откладываются питательные вещества). В эндосперме развиваются архегонии — женские органы полового размножения, в которых образуются яйцеклетки.
В период опыления из микропиле выделяется жидкость, в которую попадают пыльцевые зерна; у ели они втягиваются в пыльцевые камеры по мере высыхания жидкости (у сосны втягиванию пыльцевого зерна способствуют воздушные мешки пылинки). Когда женские шишки приняли пыльцу, их чешуи закрываются, а сами шишки постепенно меняют цвет на зеленоватый. Внутри происходит таинство оплодотворения. Пыльцевое зерно прорастает внутрь, образуя пыльцевую трубку, когда она достигает яйцеклетки, ее кончик разрывается и высвобождаются два спермия. Один оплодотворяет яйцеклетку, а второй отмирает. Из оплодотворенной яйцеклетки (зиготы) развивается зародыш, это уже спорофит (диплоидная стадия жизненного цикла). Семязачаток в это время становится семенем, шишка увеличивается в размерах и поникает на ветке. Происходит процесс одревеснения, и шишка опять меняет цвет — уже на привычный нам коричневый.
Когда женские шишки приняли пыльцу, их чешуи закрываются, а сами шишки постепенно меняют цвет на зеленоватый. Внутри происходит таинство оплодотворения. Пыльцевое зерно прорастает внутрь, образуя пыльцевую трубку, когда она достигает яйцеклетки, ее кончик разрывается и высвобождаются два спермия. Один оплодотворяет яйцеклетку, а второй отмирает. Из оплодотворенной яйцеклетки (зиготы) развивается зародыш, это уже спорофит (диплоидная стадия жизненного цикла). Семязачаток в это время становится семенем, шишка увеличивается в размерах и поникает на ветке. Происходит процесс одревеснения, и шишка опять меняет цвет — уже на привычный нам коричневый.
К осени формирование семени завершается, но ель не торопится избавиться от семян — чешуи шишек раскроются зимой или в начале весны, когда образуется наст. Это время удобно тем, что легкие крылатые семена скользят по насту и ветром перемещаются на большие расстояния. Животные (например, белки) тоже способствуют переносу, однако строение семени больше приспособлено к перемещению с помощью ветра. Когда снег тает, семена попадают в землю, а с наступлением тепла прорастают и дают начало новым елям.
Когда снег тает, семена попадают в землю, а с наступлением тепла прорастают и дают начало новым елям.
План строения и размножения у обыкновенной сосны такой же, отличаются только некоторые детали. В конце мая — начале июня у сосны формируются микростробилы в основании растущих побегов. Они напоминают пухлые колоски желто-зеленого или розового цвета. А к началу пыления уже вырастают женские шишки, они очень маленькие, до сантиметра в длину, и тоже розового цвета. Пыльца с мужских шишек попадает на мегастробилы, но до оплодотворения пройдет еще 12–14 месяцев.
Следующим летом оплодотворенная женская шишка уже поменяет свой цвет на зеленый и увеличится в размерах. Она будет висеть вбок от стебля, а над ней начнет расти новый побег. Затем шишка увеличится и станет коричневой, а дождавшись зимы, тоже рассыпет свои крылатые семена по насту.
Похожий план размножения наблюдается у множества хвойных растений. Однако у разных видов свои особенности. У тиса ягодного семена окружены красной мясистой оболочкой — ариллусом, или присемянником, — способствующей распространению с помощью птиц (см. картинку дня Ядовитый тис). У можжевельника образуются «шишкоягоды» — их чешуи сочные и сросшиеся (они разрастаются и смыкаются уже после оплодотворения). Лиственница на зиму сбрасывает всю хвою, и стробилы образуются раньше, чем вырастут новые хвоинки: ничто не мешает пыльце распространяться и попадать в женские шишки. А у сибирской сосны семена имеют плотные покровы (это те самые кедровые орешки) и приспособлены к переносу не ветром, а животными.
У тиса ягодного семена окружены красной мясистой оболочкой — ариллусом, или присемянником, — способствующей распространению с помощью птиц (см. картинку дня Ядовитый тис). У можжевельника образуются «шишкоягоды» — их чешуи сочные и сросшиеся (они разрастаются и смыкаются уже после оплодотворения). Лиственница на зиму сбрасывает всю хвою, и стробилы образуются раньше, чем вырастут новые хвоинки: ничто не мешает пыльце распространяться и попадать в женские шишки. А у сибирской сосны семена имеют плотные покровы (это те самые кедровые орешки) и приспособлены к переносу не ветром, а животными.
Фото © Яна Домбровская, 24 мая 2020 года.
Яна Домбровская
Сбережем зеленую ель
Сбережем зеленую ель
Занятие по ознакомлению с окружающим миром
Цели занятия:
1. Расширить знания детей о ели, познакомить с отличительными признаками сосны и ели.
2. Воспитывать бережное отношение к природе (к ели).
3. Развивать творческие способности, внимание, логическое мышление и речь учащихся.
Материалы и оборудование:
Иллюстрации с изображением ели в различные времена года; схемы-рисунки; гербарии веток и шишек ели и сосны; гербарии (или иллюстрации) кислицы, мхов, майника, грушанки, брусники, зимолюбки; репродукции картин А. А. Рылова, Левитана, Шишкина; рисунки детей; ширма и вырезанные герои сценки для теневого театра; запись песни о ёлочке (О. Фельцман – И.Шаферан).
Сообщение темы и целей занятия
Чтобы узнать, о чем мы будем говорить на нашем занятии, я предлагаю вам отгадать этот небольшой кроссворд.
1. С неба к нам он прилетел,
Покружился, тихо сел,
Замерзающую землю
В шубу белую одел. (Снег)
2. Был сперва водой
Потом
Прочным стал мостом. (Лед)
3. Назовите-ка, ребятки,
Месяц в этой вот загадке:
Дни его – всех дней короче,
Всех ночей длиннее ночи.
На поля и на луга
До весны легли снега.
Только месяц наш пройдет,
Мы встречаем Новый год. (Декабрь)
Догадались, о чем пойдет речь сегодня на нашем занятии? Что является неотъемлемой частью праздника Рождества и Нового года? Да, сегодня мы поговорим о ней, о красавице ели (учитель вывешивает на доску иллюстрацию).
Основная часть занятия
Ель обыкновенная – одна из основных лесообразующих пород наших лесов. Высота взрослого дерева может достигать 30-40 м. Ствол такого великана может достигать около метра в обхвате. Живет ель 250-300 лет. Встречаются отдельные деревья и в возрасте 400-500 лет.
Ель – удивительно красивое и стройное растение с тонкой островерхой кроной. Особенно хороши ели в смешанном лесу (учитель вывешивает иллюстрацию ), где их тёмная хвоя дает глубокий зелёный фон для более светлых лиственных пород деревьев. Ель требовательна к влажности и минеральному составу почвы. Активно заселяет глинистые и суглинистые почвы.
Корни у ели неглубокие. В 10-15 лет у молодой ели отмирает главный, стержневой корень, и дерево удерживают только боковые корни (рисунок).
После сильного ветра чаще на опушках можно наблюдать вывороченные с землей взрослые деревья (рисунок).
Ель – это вечнозелёное растение. Листва, а вернее хвоя, похожа на иголки. Ели сбрасывают свою хвою каждые пять-семь лет. Отжившая хвоя опадает в течение всего года. Это хорошо видно зимой: под ёлками на снегу всегда есть опавшие хвоинки. Дети рассматривают гербарии ели и сосны на столах (рисунок).
Рассмотрите ветки ели и сосны. Чем они похожи и чем различаются? У какого дерева длиннее иголки? Какие по форме шишки у сосны и у ели? Знаете ли вы, что в шишках созревают семена? Семечко сбоку имеет прозрачное пленчатое крылышко. Отличить семена ели от семян сосны на первый взгляд непросто. Но если отделить само семя ели от крылышка, то на крылышке останется ямка, а у сосны – сквозное отверстие (рисунок).
Ели не страшен любой снегопад. Под тяжестью снега ветви ее пригибаются, и снег сползает с них. Широкие ветви ели (“лапы”) очень пружинистые. Снег гнёт их к земле, но никогда не ломает (иллюстрации).
Под тяжестью снега ветви ее пригибаются, и снег сползает с них. Широкие ветви ели (“лапы”) очень пружинистые. Снег гнёт их к земле, но никогда не ломает (иллюстрации).
В 35-45 лет начинает плодоносить. Созревшие семена высыпаются из шишек во второй половине зимы. Ветер подхватывает эти “парусники” и гонит их по гладкому насту – так ель расселяется.
До 10-15 лет ель растет очень медленно, затем рост усиливается. Ель растет всю свою жизнь, поэтому у неё всегда острая вершина. Верхушка ели – это однолетний вертикальный побег, в основании которого заложено кольцо почек, из которых на следующий год вырастут боковые ветки, образующие мутовку (рисунок).
Ель обыкновенная не боится затенения и хорошо растёт под пологом других растений. Это дерево исключительно морозостойкое, однако молодые ёлочки очень чувствительны к поздним весенним заморозкам. Весной ели трогаются в рост. Почки развёртываются, образуя щепочки нежной светло-зелёной хвои на молодом побеге. Но стоит ударить весенним заморозкам, как этот прирост будет уничтожен. А под пологом смешанного или лиственного леса ели защищены от губительного действия этих морозов.
А под пологом смешанного или лиственного леса ели защищены от губительного действия этих морозов.
Травянистый покров ельника беден. В ельнике образуется толстая подстилка, препятствующая распространению травянистых растений. Почва под такой подстилкой зимой сильно промерзает, а весной снег сходит очень поздно, так как почва в тени долго не прогревается.
Для травянистого покрова ельника характерны различные виды мхов, кислица обыкновенная, майник двулистный и др., причем половина спутников ели – грушанка, брусника, зимолюбка – зимует с зелеными листьями, так же как и сама ель (речь учителя сопровождается иллюстрациями или демонстрацией гербария).
Сделай вывод:
1. Дерево имеет только один крепкий, часто высокий древесный стебель – ствол. Ель имеет один ствол. Значит ель – ….
2. Все хвойные растения имеют хвою. У ели листья видоизменены в хвоинки. Значит ель – ….
3. У теневыносливых растений ветви растут до земли. У ели густая конусовидная крона. Значит ель – …
А теперь послушайте экологический рассказ-загадку, который называется “Зимний день”. Ваша задача – внимательно его послушать и найти ошибки.
Ваша задача – внимательно его послушать и найти ошибки.
Зимний день
В классе только и было разговоров о том, будет в воскресенье мороз или оттепель. Проснулись утром и первым делом посмотрели на термометр: за окном – 4 градуса мороза. Едем! Поезд привез нас к лесу часов в девять. Тихо зимой в лесу, и только кое-где слышны голоса синиц и скворцов. Накануне выпал снег, и хорошо было видно следы некоторых жителей леса. Вот пробежал заяц. А это чьи следы подле просеки? “Это, наверное, следы ежа”, – сказал кто-то из ребят. Неожиданно мы вышли к опушке, где росли одинокие ели. На белоснежной картине выделялись несколько огромных темных стволов, которые оканчивались высокими кронами. Иглы деревьев уже давно пожелтели и постепенно опадали на снег. Постоянно слышался треск сучьев. Мы подняли головы вверх. Теперь стало всё ясно. Под тяжестью выпавшего снега ветви елей трещали, ломались и падали вниз. На ветке одной густой ели мы увидели гнездо и в нём какую-то птицу. Кто же это насиживает яйца зимой? Это сойка! Солнце уже приближалось к горизонту, и мы двинулись к станции. Подходя к полю, мы увидели много звериных следов возле стога сена. “Наверное, это волк ловил мышей в сене”, – подумали дети. Через несколько минут поезд уже набирал скорость. Немного усталые, но довольные, мы подъезжали к городу.
Подходя к полю, мы увидели много звериных следов возле стога сена. “Наверное, это волк ловил мышей в сене”, – подумали дети. Через несколько минут поезд уже набирал скорость. Немного усталые, но довольные, мы подъезжали к городу.
Ошибки:
1. Скворцы – перелётные птицы, зимой их не бывает.
2. Ежи зимой впадают в спячку.
3. Крона у ели конусовидная, а ветви растут до самой земли.
4. Хвоя у елей не желтеет и опадает постепенно в течение года.
5. Под тяжестью снега ветви ели не ломаются, а пригибаются и снег сползает.
6. Зимой насиживает яйца не сойка, а клёст.
7. “Мышкует” в сене не волк, а лиса.
Ель очень полезна для человека. Древесина её мягкая и лёгкая. Она используется в строительстве, в целлюлозно-бумажной промышленности, незаменима при изготовлении музыкальных инструментов (это так называемая резонансная ель, годичные кольца которой имеют одинаковую ширину). Из еловой древесины вырабатывают деготь, канифоль, скипидар, смолу. Хвоя служит источником витамина С, из коры получают дубильные вещества.
Хвоя служит источником витамина С, из коры получают дубильные вещества.
Существуют и народные приметы, связанные с елью. Давайте вспомним знакомые и узнаем новые. (Нужно соединить стрелочками первую и второю половину приметы).
1. Если ветви ели опускаются вниз – к оттепели.
2. Еловый лес зимой чернеет – к дождю.
3. Если в безветренную погоду сухие ветви падают с деревьев – перед ясной погодой.
4. Если ветви ели поднимаются вверх – к дождю.
5. Если шишки на ели растут низко – к холодам в конце зимы.
6. Если шишки на ели растут высоко – к ранним морозам.
Кроме этого ель имеет огромное эстетическое значение для человека. Красота этого дерева вдохновляла многих поэтов, художников и музыкантов. Вот послушайте стихотворения, которые приготовили для вас ваши товарищи.
Вырастала ёлка в лесу на горе,
У неё иголки зимой в серебре,
У неё на шишках ледышки стучат,
Снежное пальтишко лежит на плечах…
(Е. Трутнева)
Ни листочка, ни травинки!
Тихим стал наш сад.
И берёзки и осинки
Скучные стоят.
Только ёлочка одна
Весела и зелена.
Видно, ей мороз не страшен,
Видно, смелая она!
(О. Выготская)
Ели на опушке – до небес макушки –
Слушают, молчат, смотрят на внучат.
А внучата-ёлочки, тонкие иголочки –
У лесных ворот водят хоровод.
(И. Токмакова)
А теперь посмотрите на эти репродукции. Перед вами репродукция картины А. А. Рылова “Тайга”. Огромные деревья увязли в глубоких сугробах. Их ветви гнутся под тяжёлыми шапками снега. Высокие ели стоят плотной стеной, отбрасывая на снег синие тени. Это же чудо! А как красивы ели на репродукциях картин Левитана и Шишкина!?
Вы тоже готовились к этому занятию и ёлочки на ваших рисунках тоже очень хороши. А я хочу обратить ваше внимание на рисунки, где изображены украшенные различными игрушками и сладостями ели. Атрибутом каких праздников является такая ель? А знаете ли вы, почему в Рождество и Новый год принято наряжать именно это дерево? Давайте посмотрим спектакль. (Теневой театр).
(Теневой театр).
(Выходит девочка, наряженная ёлочкой)
Ёлочка:
Ясная ночь. Тихо кругом.
Ярко горит звезда над пещерой.
Ангелов хор смолк за холмом.
Свет голубой льётся из щелей.
В яслях Младенец-Спаситель лежит,
Тысячи лет Его ждали прихода.
Счастлив тот будет, кто к Нему поспешит.
Счастливы люди! Ликует природа!
(пробегает Зайчик)
Зайчик:
Скок-поскок,
Путь недалёк.
С дороги не сбиться –
Вижу звезду.
Спешу поклониться
Младенцу-Христу.
Ёлочка:
Беги, Заинька, беги.
Зайчик:
Но у меня ничего нет подарить Ему!
Ёлочка:
Ничего, Заинька. У тебя такие мягкие ушки – Младенец погладит их, и Ему будет весело.
Зайчик:
Правда? Вот спасибо, Ёлочка.
Скок-поскок,
Путь недалёк…
Ёлочка:
Беги, Зайчик!
Беги, маленький!
(Выходит медведь, напевая)
Медведь:
А я Мишка-медведь,
Я пришёл поглядеть
На Божественного Младенца.
Ой, а что мне подарить ему?
Ёлочка:
А ты покувыркайся перед Ним, Миша, ты ведь такой ловкий. Младенец очень обрадуется.
Младенец очень обрадуется.
Медведь:
Вот спасибо, Ёлочка, что научила. Побегу скорее.
(Вбегают цветы, кружатся, поют)
Цветы:
Мы цветы-цветочки,
Мы растём неслышно. Эта ночка-ноченька
Самая душистая.
Ёлочка:
Куда это вы, Цветы? Вам спать ночью полагается…
Цветы:
Это ночка-ноченька
Света ярче светится.
Поклониться в ноженьки
Мы идём Младенцу.
Ёлочка:
Возьмите и меня с собой, Цветы мои милые, возьмите и меня поклониться Христу-Младенцу.
Цветы:
Но на тебе нет цветков, Ёлочка, а о твои иголки Младенец может только уколоться. (Уходят).
Ёлочка:
Никого. Одна я, одинока.
Ночь тиха. Звезда зовёт, горит.
Все ушли… Все поклониться Богу
К святой пещере радостно спешат.
Колючая, забытая ёлка,
Ненужная стою в своей глуши.
Цветы правы. Я за Младенца
Здесь помолюсь тихонько от души.
(Возвращаются цветы)
1 цветок:
Ах, Ёлочка, Пальма положила Младенцу к ногам свою лучшую ветвь и сказала: “Пусть она навевает на Тебя прохладу в жаркий день”.
2 цветок:
А маслина наклонила свои ветви, и с них закапало душистое масло.
3 цветок:
И вся пещера наполнилась благоуханием.
Ёлочка:
А я такая бедная и ничтожная. Мне нечем порадовать Младенца…
Появляется ангел.
Ангел:
Не грусти, Ёлочка! За твою доброту и скромность я украшу тебя лучше твоих сестёр. Я украшу тебя звёздами, и ты порадуешь Младенца. Доброе деревце, отныне, по воле Божьей, ты будешь каждый год красоваться в сиянии огней, и маленькие дети, глядя на тебя, будут радоваться и веселиться. И ты, скромная зелёная ёлка. Станешь знамением весёлого рождественского праздника!
Праздник – это радость, смех, веселье. Но всем ли радостно в новогоднюю ночь?! Вот послушайте стихотворение, которое написала О. Беляевская.
Ёлка зелёная, густоветвистая
В пышный наряд убрана;
В пышном уборе роняет смолистые
Слёзы беззвучно она.
Ноет в стволе её рана глубокая,
Сердце ей точит печаль:
Леса родимого, леса далёкого
Ёлке мучительно жаль.
Звёздного неба пространство безбрежное
Там возвышалось над ней;
Стлались под нею ковры белоснежные
С синим узором теней.
Ветер ей пел свои песни свободные;
Сказки ей лес говорил;
Щедрый мороз её ночью холодною
В жемчуг, в алмазы рядил.
Думает ёлка с тоскою глубокою:
“Лес мой родной, не видать
Мне тебя больше. Прости! Одинокою
Здесь суждено умирать.
А для чего?!”
А вкруг ёлки шумливою
Дети собрались толпой.
Дышат весельем их лица счастливые,
Радостью блещут живой.
Ёлка задумалась. Утро весеннее.
Лепет зелёных ветвей,
Птиц на заре щебетанье и пение
Дети напомнили ей.
Радость их горе смягчила тяжёлое.
Тайной отрадой полна,
Вспыхнула ярко огнями весёлыми
В грустной неволе она.
Кто же с грустью встречает праздник? Почему ёлочке грустно? Вы только представьте, сколько нужно срубить ёлочек, чтобы встретить Новый год по традиции каждой семье!!!! И это повторяется каждый год. Если не остановить этот губительный процесс, то наступит время, когда на нашей планете не останется ни одной ели!!! Но это ещё не всё… (Учитель вывешивает на доску рисунок).
Глядя на рисунок, представь, что ты и сам очутился в этом лесу, рядом с огромной елью. Будь осторожен, чтобы не спугнуть лесных обитателей. Смотри-ка, одни животные питаются семенами еловых шишек, живут на ели. Эти животные напрямую связаны с елью. А на них, на этих животных, охотятся другие. Значит, и они тоже с елью связаны, только не напрямую, а, как, говорят учёные, косвенно. Если люди срубят ель, плохо придётся лесным животным. Не только растительноядным, но и насекомоядным, и хищникам! Объясните – почему?
Что же должны делать люди, чтобы на долгие тысячелетия сохранить красавицу ель?
(Ученик читает стихотворение)
В лесу родилась ёлочка,
В лесу должна расти.
Не нужно рубить ёлочку.
И в дом её нести.
В лесу так ёлке весело,
Метель поёт ей песенки.
Укрыты веточки снежком,
Стоят все ёлочки рядком.
Не мёрзнет ёлочка зимой
Не просит брать её домой.
Идите в лес вы к ёлке сами,
И праздник встретьте там с друзьями.
Не стоит ёлочку губить,
А лучше жизнь ей сохранить.
Искусственную ёлочку купите,
И в своём доме нарядите.
Итак, как можно встретить Рождество и Новый год и не погубить ёлочку? Купить искусственную ёлку, посадить ёлочку во дворе и украсить её, организовать зимнюю прогулку в лес к ёлочке, украсить еловые веточки, сделать ёлочки из цветной бумаги на уроке труда.
И всё же давайте не будем забывать о том, что в праздники принято веселиться и играть. Давайте и мы на нашем занятии немного поиграем.
Отгадайте ребусы
(щавель, капель, портфель, пчёлка, метель, газель, учитель, кисель, китель, кашель)
Давайте послушаем песенку о ёлочке. Кто знает слова, может подпевать.
Подведение итогов
Что узнали нового? Какие правила вывели для себя?
И последнее задание – мысленно поменяйте одинаковые ёлочки местами, и вы узнаете, как вы сегодня работали (МОЛОДЦЫ!).
Литература:
1. Начальная школа, №5.-1990
2. Я. С. Хренов.- Народные приметы и календарь.-1991.-Москва
Хренов.- Народные приметы и календарь.-1991.-Москва
3. Начальная школа, №4.- 1992
4. Праздник Рождества Христова.- Сборник материалов для организации праздника.- Москва.- 2000
5. Серебряный звон: Учебное пособие./ Сост., предисл., примеч. свящ. Д. Лескин. – Тольятти, 1999
© Все права защищены http://www.portal-slovo.ru
Кисловодская ель – претендент на победу в конкурсе «Российское дерево года»
+ A –
Это дерево представляет собой культурную, историческую и эстетическую ценность
Первого мая на сайте www.rosdrevo.ru стартовал конкурс «Российское дерево года», который продлится до первого августа. В списке номинантов под №4 для голосования значится «Ель обыкновенная», которая произрастает на территории литературно-музыкального музея «Дача Шаляпина» в Кисловодске.
Это дерево представляет собой культурную, историческую и эстетическую ценность, кисловодская ель внесена в Национальный реестр старовозрастных деревьев России под номером анкеты 284.
Организатор российского конкурса – НПСА «ЗДОРОВЫЙ ЛЕС». Одна из задач конкурса – вдохновить активных граждан заботиться об окружающей среде, обратить их внимание на значение старовозрастных деревьев в природном и культурном наследии России, которое требует нашей заботы и защиты. Победитель номинируется к участию в международном конкурсе «Европейское дерево года-2022». Представителю органа исполнительной власти субъекта РФ, на территории которого произрастает дерево, вручается Диплом победителя, а также Сертификат на обследование дерева и проведение оздоровительных мероприятий специалистами Центра древесных экспертиз «ЗДОРОВЫЙ ЛЕС».
В рамках российского этапа, на территории региона, где произрастает дерево-победитель, проводится конкурс детского рисунка. Работы победителей отправляются в Брюссель для участия в выставке, которая проходит в здании Европейского Парламента во время церемонии объявления итогов международного конкурса «Европейское дерево года».
Министерство природных ресурсов и охраны окружающей среды СК призывает жителей Ставропольского края проголосовать за кисловодскую ель в конкурсе «Российское дерево года-2021» на сайте www.rosdrevo.ru.
– Старовозрастные деревья – уникальные представители растительного мира, живые свидетели исторических событий, связующее звено ушедших и будущих поколений, наше национальное достояние. От вашего выбора зависит, какое дерево будет представлять нашу страну на европейском соревновании, – подчеркнул и. о. министра природных ресурсов и охраны окружающей среды Ставропольского края Андрей Хлопянов.
Опубликован в газете “Московский комсомолец” №23 от 2 июня 2021
Заголовок в газете: Выбрать хвойное до августа
Ель обыкновенная: многоцелевое дерево
Picea abies
Англичане не стали называть норвежскую ель по имени, вместо этого называя ее «ель обыкновенная». Финны считали его своим, называя финской елью, а другие – европейской елью. Независимо от того, как вы это называете, европейская ель является основным продуктом питания в Европе. Ель обыкновенная, наиболее известная своей прочностью и высокой высотой, распространила свою популярность через Атлантику и в США.С., став американским фаворитом.
Финны считали его своим, называя финской елью, а другие – европейской елью. Независимо от того, как вы это называете, европейская ель является основным продуктом питания в Европе. Ель обыкновенная, наиболее известная своей прочностью и высокой высотой, распространила свою популярность через Атлантику и в США.С., став американским фаворитом.
Что делает это дерево еще более привлекательным, так это его многократное использование. Это важная древесная культура в Европе, позволяющая производить прочную легкую древесину с прямыми волокнами, что делает ее идеальным выбором для строительства. Это отличное ландшафтное дерево с густой листвой и высокой высотой. Естественная пирамидальная форма и зеленый цвет дерева делают его одной из самых популярных рождественских елок в стране.
Вот несколько моментов, на которые следует обратить внимание, если вы планируете добавить ель обыкновенную к своей древесной семье.
Условия окружающей среды:
- Хорошо проявляет себя на кислых, глинистых, суглинистых, влажных, песчаных и хорошо дренированных почвах (зоны зимостойкости 3-7).
- Хорошо работает как на полном, так и на частичном солнце.
- Дерево от среднего до быстрорастущего, вырастающее до двух футов в год и достигающее 40-60 футов в зрелом возрасте.

Физические атрибуты:
- Имеет темно-зеленые, дюймовые иглы с квадратными кончиками, иглы сохраняются от шести до семи лет до опадания.
- Имеет тонкую красновато-коричневую кору, которая с возрастом утолщается и отслаивается.
- Шишки начинают формироваться в возрасте 30 лет, при этом семена опадают зимой или ранней весной, обеспечивая пищу для диких животных.
Отметьте нас на фото со своей елью обыкновенной!
Ширин Осман
Сотрудник по коммуникациям
Ель обыкновенная с признаками высыхания (посередине) среди здоровых…
Контекст 1
… Считается, что на юге Норвегии верхнее усыхание древостоев ели обыкновенной (Picea abies (L. ) Karst.) Возрастом 40-50 лет связано с экстремальными климатическими условиями ( Hentschel et al.2014). В этом регионе период с 2004 по 2006 год характеризовался высоким дефицитом воды в течение вегетационного периода, который оценивался как разница между кумулятивными осадками и потенциальной эвапотранспирацией (см. Рис.1 в Hentschel et al.2014). В этот период деревья с симптомами высыхания верхушек (рис.1) показали более низкую внутреннюю эффективность использования воды, которая связана с повышенной устьичной проводимостью (Hentschel et al., 2014), и давали древесину с меньшей плотностью, что указывает на более высокую уязвимость к кавитации (Rosner и др., 2014). Функциональные характеристики для оценки гидравлической уязвимости приобретают все большее значение в качестве инструментов скрининга, поскольку гидравлические эксперименты трудоемки или подвержены ошибкам (Cochard et al. 2013). В этом исследовании мы расширяем существующий набор данных (Rosner et al.2014) с образцами деревьев и получить функциональные характеристики гидравлической уязвимости на основе размеров трахеид, оцененных с помощью технологии SilviScan (Evans 1994).
) Karst.) Возрастом 40-50 лет связано с экстремальными климатическими условиями ( Hentschel et al.2014). В этом регионе период с 2004 по 2006 год характеризовался высоким дефицитом воды в течение вегетационного периода, который оценивался как разница между кумулятивными осадками и потенциальной эвапотранспирацией (см. Рис.1 в Hentschel et al.2014). В этот период деревья с симптомами высыхания верхушек (рис.1) показали более низкую внутреннюю эффективность использования воды, которая связана с повышенной устьичной проводимостью (Hentschel et al., 2014), и давали древесину с меньшей плотностью, что указывает на более высокую уязвимость к кавитации (Rosner и др., 2014). Функциональные характеристики для оценки гидравлической уязвимости приобретают все большее значение в качестве инструментов скрининга, поскольку гидравлические эксперименты трудоемки или подвержены ошибкам (Cochard et al. 2013). В этом исследовании мы расширяем существующий набор данных (Rosner et al.2014) с образцами деревьев и получить функциональные характеристики гидравлической уязвимости на основе размеров трахеид, оцененных с помощью технологии SilviScan (Evans 1994). После этого мы документируем, как эти прокси изменились с годами у 20 здоровых и 20 падающих деревьев, чтобы узнать больше о факторах, предрасполагающих к отмиранию верхушки ели норвежской на юге Норвегии. Десять деревьев здорового вида и десять деревьев с визуальными признаками отмирания верхушек, такими как сухие верхушки и пожелтение хвои в …
После этого мы документируем, как эти прокси изменились с годами у 20 здоровых и 20 падающих деревьев, чтобы узнать больше о факторах, предрасполагающих к отмиранию верхушки ели норвежской на юге Норвегии. Десять деревьев здорового вида и десять деревьев с визуальными признаками отмирания верхушек, такими как сухие верхушки и пожелтение хвои в …
Контекст 2
… южная часть Норвегии, недавнее высыхание верхушки насаждений ели европейской (Picea abies (L.) Karst.) возрастом 40-50 лет, как полагают, связано с экстремальными климатическими условиями (Hentschel et al. 2014). В этом регионе период с 2004 по 2006 год характеризовался высоким дефицитом воды в течение вегетационного периода, который оценивался как разница между совокупным количеством осадков и потенциальной эвапотранспирацией (см. Рис. 1 в Hentschel et al. 2014). В этот период деревья с признаками отмирания верхушки (рис.1) показали более низкую внутреннюю эффективность водопользования, связанную с повышенной проводимостью устьиц (Hentschel et al. 2014), и производили древесину с более низкой плотностью, что указывает на более высокую уязвимость к кавитации (Rosner et al. 2014). Функциональные характеристики для оценки гидравлической уязвимости приобретают все большее значение в качестве инструментов скрининга, поскольку гидравлические эксперименты трудоемки или подвержены ошибкам (Cochard et al. 2013). В этом исследовании мы расширяем существующий набор данных (Rosner et al. 2014) образцами деревьев и выводим функциональные характеристики гидравлической уязвимости на основе размеров трахеид, оцененных с помощью технологии SilviScan (Evans 1994).После этого мы документируем, как эти прокси изменились с годами у 20 здоровых и 20 падающих деревьев, чтобы узнать больше о факторах, предрасполагающих к отмиранию верхушки ели норвежской на юге Норвегии. Десять здоровых деревьев и десять деревьев с визуальными признаками отмирания верхушек, такими как сухие верхушки и пожелтение хвои в …
2014), и производили древесину с более низкой плотностью, что указывает на более высокую уязвимость к кавитации (Rosner et al. 2014). Функциональные характеристики для оценки гидравлической уязвимости приобретают все большее значение в качестве инструментов скрининга, поскольку гидравлические эксперименты трудоемки или подвержены ошибкам (Cochard et al. 2013). В этом исследовании мы расширяем существующий набор данных (Rosner et al. 2014) образцами деревьев и выводим функциональные характеристики гидравлической уязвимости на основе размеров трахеид, оцененных с помощью технологии SilviScan (Evans 1994).После этого мы документируем, как эти прокси изменились с годами у 20 здоровых и 20 падающих деревьев, чтобы узнать больше о факторах, предрасполагающих к отмиранию верхушки ели норвежской на юге Норвегии. Десять здоровых деревьев и десять деревьев с визуальными признаками отмирания верхушек, такими как сухие верхушки и пожелтение хвои в …
Frontiers | Функциональные отношения анатомических признаков древесины у европейской ели
Введение
Каждая отдельная клетка в кольце дерева происходит от деления камбиальной клетки (периклинальный рост), которая затем расширяется до своего окончательного размера, утолщает свою клеточную стенку и в конечном итоге лигнифицируется до запрограммированной гибели клеток (Plomion et al. , 2001). Эти шаги запускаются и управляются сложным взаимодействием генетических факторов, онтогенеза (развития организма с течением времени), регуляции гормонов и условий окружающей среды (Aloni, 2013). Формирование древесных клеток, таким образом, представляет собой уникальную и сложную комбинацию взаимодействующих процессов, приводящих к полному древовидному кольцу (Vaganov et al., 2006; Fonti and Jansen, 2012). Обширная литература посвящена задаче связи клеточной структуры древесины с ее функционированием (например, Tyree and Ewers, 1991; Domec et al., 2009; Фонти и др., 2013; Hacke et al., 2015; Бекман, 2016; Де Микко и др., 2019). Например, было продемонстрировано, что диаметр и плотность трахеид, процентное соотношение ранней и поздней древесины и толщина клеточной стенки (CWT) играют центральную роль в регулировании гидравлической эффективности и механического функционирования дерева (например, McCulloh et al., 2010; Rosner, 2013; Prendin et al., 2018), но также сильно зависит от климата (например, Larson, 1994; Carrer et al.
, 2001). Эти шаги запускаются и управляются сложным взаимодействием генетических факторов, онтогенеза (развития организма с течением времени), регуляции гормонов и условий окружающей среды (Aloni, 2013). Формирование древесных клеток, таким образом, представляет собой уникальную и сложную комбинацию взаимодействующих процессов, приводящих к полному древовидному кольцу (Vaganov et al., 2006; Fonti and Jansen, 2012). Обширная литература посвящена задаче связи клеточной структуры древесины с ее функционированием (например, Tyree and Ewers, 1991; Domec et al., 2009; Фонти и др., 2013; Hacke et al., 2015; Бекман, 2016; Де Микко и др., 2019). Например, было продемонстрировано, что диаметр и плотность трахеид, процентное соотношение ранней и поздней древесины и толщина клеточной стенки (CWT) играют центральную роль в регулировании гидравлической эффективности и механического функционирования дерева (например, McCulloh et al., 2010; Rosner, 2013; Prendin et al., 2018), но также сильно зависит от климата (например, Larson, 1994; Carrer et al. , 2016; Castagneri et al., 2017). Иными словами, на некоторые гидравлические параметры, такие как гидравлический диаметр и удельную гидравлическую проводимость, влияет высота дерева (например,г., Анфодилло и др., 2013; Rosell et al., 2017). Однако межличностные и внутрииндивидуальные вариации нескольких анатомических черт древесины редко анализировались на большом количестве деревьев, растущих в разных условиях окружающей среды (Rungwattana and Hietz, 2017; Prendin et al., 2018). Как следствие, до сих пор неясно, насколько значимым является вклад каждого фактора, уравновешивающего компромисс в клетке древесины и, следовательно, на уровне годичных колец. Дендрофенотипические признаки (например, Evans et al., 2018; Heer et al., 2018), а также разработка оптимальных стратегий для количественной оценки их вариабельности важны для оценки влияния потенциальных факторов индивидуальной вариабельности в чертах роста. Недавно было проведено несколько исследований, которые позволили по-новому взглянуть на межиндивидуальные вариации анатомических характеристик древесины, подчеркнув роль высоты дерева как движущей силы анатомических вариаций древесины вдоль стебля растения, в отличие от классической тенденции возраста от сердцевины до коры (Enquist, 2002; Olson et al.
, 2016; Castagneri et al., 2017). Иными словами, на некоторые гидравлические параметры, такие как гидравлический диаметр и удельную гидравлическую проводимость, влияет высота дерева (например,г., Анфодилло и др., 2013; Rosell et al., 2017). Однако межличностные и внутрииндивидуальные вариации нескольких анатомических черт древесины редко анализировались на большом количестве деревьев, растущих в разных условиях окружающей среды (Rungwattana and Hietz, 2017; Prendin et al., 2018). Как следствие, до сих пор неясно, насколько значимым является вклад каждого фактора, уравновешивающего компромисс в клетке древесины и, следовательно, на уровне годичных колец. Дендрофенотипические признаки (например, Evans et al., 2018; Heer et al., 2018), а также разработка оптимальных стратегий для количественной оценки их вариабельности важны для оценки влияния потенциальных факторов индивидуальной вариабельности в чертах роста. Недавно было проведено несколько исследований, которые позволили по-новому взглянуть на межиндивидуальные вариации анатомических характеристик древесины, подчеркнув роль высоты дерева как движущей силы анатомических вариаций древесины вдоль стебля растения, в отличие от классической тенденции возраста от сердцевины до коры (Enquist, 2002; Olson et al. , 2014; Kašpar et al., 2019).Это особенно верно для каналов у древесных пород, предназначенных для транспортировки воды от корней к листьям. Согласно гидродинамике, увеличение высоты растений будет сопровождаться одновременным увеличением гидродинамического сопротивления вдоль канала (West et al., 1997). Этот эффект может ограничивать рост растения в высоту, если он не компенсируется увеличением размеров древесных каналов во время онтогенеза. Неоднократно наблюдалось, что диаметр канала становится больше (и длиннее) по направлению к основанию растения по мере увеличения расстояния до верхушки дерева, что называется «расширением канала» (Anfodillo et al., 2013; Lazzarin et al., 2016; Rosell et al., 2017). Расширение канала, если смотреть на поперечный разрез древесины от основания ствола, представлено увеличением размера канала от сердцевины до коры (как описано Malpighi, 1675), и это часто интерпретируется как возрастная тенденция. (Gartner, 1995; Lachenbruch et al., 2011). В короткой установке с узкими трубопроводами увеличение размера канала от сердцевины до коры является следствием того, что каналы становятся больше, чтобы компенсировать увеличение высоты (Carrer et al., 2015) (рисунок 1).
, 2014; Kašpar et al., 2019).Это особенно верно для каналов у древесных пород, предназначенных для транспортировки воды от корней к листьям. Согласно гидродинамике, увеличение высоты растений будет сопровождаться одновременным увеличением гидродинамического сопротивления вдоль канала (West et al., 1997). Этот эффект может ограничивать рост растения в высоту, если он не компенсируется увеличением размеров древесных каналов во время онтогенеза. Неоднократно наблюдалось, что диаметр канала становится больше (и длиннее) по направлению к основанию растения по мере увеличения расстояния до верхушки дерева, что называется «расширением канала» (Anfodillo et al., 2013; Lazzarin et al., 2016; Rosell et al., 2017). Расширение канала, если смотреть на поперечный разрез древесины от основания ствола, представлено увеличением размера канала от сердцевины до коры (как описано Malpighi, 1675), и это часто интерпретируется как возрастная тенденция. (Gartner, 1995; Lachenbruch et al., 2011). В короткой установке с узкими трубопроводами увеличение размера канала от сердцевины до коры является следствием того, что каналы становятся больше, чтобы компенсировать увеличение высоты (Carrer et al., 2015) (рисунок 1).
Рис. 1. Эскиз масштабирования структуры древесины от сердцевины до коры в зависимости от высоты дерева (модифицированный из Rosell et al., 2017) на гипотетическом 5-летнем дереве, предполагая тот же радиальный (древовидный) ширина кольца) и продольный (длина междоузлия) рост. Показано изменение количества и размера трахеид от сердцевины к коре. У коротких деревьев (или близко к сердцевине) больше трахеид меньшего размера (или близко к сердцевине) и меньше и более широких трахеид у высоких деревьев (или близко к коре) по сравнению с ранее сформированным кольцом.
Чтобы исследовать синергию высоты дерева, возраста и условий произрастания в окружающей среде на анатомические особенности древесины, мы количественно оценили анатомические вариации древесины в поперечных сечениях ели европейской, породы хвойных пород с простой структурой древесины, в которой трахеиды выполняют как проводящие, так и механические функции (Evert , 2006). Цели данной статьи – исследовать (1) как анатомия трахеиды в радиальном и тангенциальном направлениях связана с онтогенезом, (2) как анатомические вариации древесины отражают функциональные ограничения и компромиссы, и (3) как анатомические черты древесины меняются в зависимости от межиндивидуальный и межгодовой уровень.Чтобы свести к минимуму влияние климата на анатомические характеристики древесины, которые в данном исследовании рассматриваются как «шум», были проанализированы только пять внешних годичных колец (2008–2012 гг.). Результаты предоставили сводную информацию об изменчивости анатомических признаков древесины и предложили возможные последствия для связанных областей исследований, таких как экология растений и эволюционная генетика.
Материалы и методы
Место исследования и сбор образцов
Полевой отбор проб проводился в 2012 г. в природном заповеднике Камполино (северные Апеннины) (таблица 1), который является самой южной известной автохтонной популяцией ели европейской [ Picea abies (L.) H. Karst.] В Италии (Chiarugi, 1936; Magri et al., 2015; Avanzi et al., 2019). Вдоль высотного разреза на одном склоне заложены три участка. Высокогорный участок (CAM-H, ~ 1730 м над ур. М.) Представляет собой чистый, густой и относительно молодой еловый лес, переселяющий старые пастбища. Промежуточный участок (CAM-E, ∼1615 м над ур. Самый низкий участок (CAM-L, ∼1,475 м над уровнем моря).s.l.) – аналогичный смешанный лес, но с меньшим количеством елей (таблица 1). На каждом выбранном участке деревья были помечены и привязаны к местности с помощью устройства системы географического позиционирования (GPS; Garmin, разрешение 2 м). Диаметр ствола на высоте груди (DBH) и высота дерева были измерены, и один керн был собран с помощью 5-миллиметрового сверлильного станка Pressler на DBH в поперечном направлении, чтобы избежать сжатия древесины.
Таблица 1. Географические координаты и характеристики древостоя трех елей европейской [ Picea abies (L.) Х. Карст.] Сюжеты.
Три участка попадают в одну и ту же пространственную сетку, поэтому среднемесячные температуры и общее количество осадков за период 2008–2012 гг. Были получены из сетки CRU TS V.4.0 (суша) 0,5 ° (KNMI Climate Explorer).
Выбор дерева и анатомические особенности древесины
Всего было отобрано 10 деревьев на участок (таблица 1, дополнительный рисунок S1), чтобы максимизировать вариации по высоте, возрасту и DBH деревьев (дополнительный рисунок S2). Были подготовлены микрошлифы древесины для получения анатомических изображений с высоким разрешением в соответствии со стандартными протоколами (Gärtner and Schweingruber, 2013; von Arx et al., 2016; Прендин и др., 2017). Из каждой керна с помощью лабораторного микротома WSL были вырезаны поперечные срезы толщиной 12–15 мкм из последних пяти годичных колец (датированные 2008–2012 гг.) (Gärtner et al., 2015a, b). Все срезы дважды окрашивали сафранином и Astra blue, обезвоживали этанолом и постоянно фиксировали на предметном стекле микроскопа канадским бальзамом. Перекрывающиеся изображения поперечных сечений получали при 100-кратном увеличении с помощью цифровой камеры (Canon EOS 650D, Canon Inc., Токио, Япония) через микроскоп Olympus BX41 (Olympus Corp., Токио, Япония). PTGUI v10 (New House Internet Services B.V., Роттердам, Нидерланды, США) использовался для сшивания отдельных изображений и создания составного изображения высокого разрешения (2,36 пикселей / мкм). Затем эти изображения были обработаны с помощью программного обеспечения ROXAS (v3.0.88, von Arx and Carrer, 2014), которое обеспечивает большой набор измерений ширины годичных колец (TRW) и анатомических характеристик древесины (Таблица 2). Анализ изображений и сбор анатомических данных были ограничены шириной одного тангенциального миллиметра в пределах каждого кольца дерева.
Таблица 2. Список всех анатомических признаков древесины, использованных в данном исследовании, с разделением по их функциям.
Анатомические признаки древесины, использованные в этом исследовании, были получены при прямых измерениях на основе результатов ROXAS (таблица 2). Анализ проводился в пределах годичного кольца деревьев, а также в ранней (EW) и поздней (LW) древесине. Были измерены TRW, ширина EW и LW (EWW и LWW) и их процентное соотношение (EW% и LW%), рассчитанные как отношение EWW (LWW) к TRW, а также их соотношение (LW / EW).Чтобы различать трахеиды EW и LW, ROXAS автоматически вычисляет индекс Морка, который представляет собой соотношение между удвоенным двойным CWT и диаметром просвета, измеренными в радиальном направлении (Denne, 1988) (Таблица 2). Кроме того, соотношение между радиальным и тангенциальным диаметром просвета трахеиды применялось для различения формы трахеиды EW (отношение> 1) и LW (отношение <1). Количество трахеид в тангенциальном направлении подсчитывалось вручную в LW, а количество трахеид в радиальном направлении вычислялось путем деления общего количества трахеид в каждом кольце дерева (выход ROXAS) на количество трахеид в тангенциальном направлении в пределах полосы 1 мм.Анатомические признаки древесины были сгруппированы по признакам, связанным с (i) ростом годичных колец, (ii) ростом клеток, (iii) гидравлической эффективностью и гидравлической безопасностью, и (iv) механической опорой и вложением углерода (Таблица 2).
Статистический анализ
Одномерный и многомерный анализ анатомических признаков древесины
Для оценки изменчивости анатомических признаков древесины, их приписываемых функций и онтогенетического эффекта был проведен одномерный и многомерный анализ.Для оценки парных отношений между анатомическими признаками древесины и влияния высоты дерева и возраста дерева были подобраны линейные, экспоненциальные и логарифмические функции, и была выбрана функция с наивысшим значением R 2 . R 2 логарифмической и экспоненциальной регрессий были рассчитаны путем логарифмического преобразования осей x и y соответственно. Анализ главных компонентов (PCA) проводился для каждого набора данных за один год с использованием функции prcomp в R (R Core Team, 2018) для одновременного описания взаимосвязей между 65 оцененными анатомическими признаками.
Определение основных источников анатомических изменений древесины
Влияние высоты, возраста и участка дерева (последний используется в качестве прокси для изменения окружающей среды вдоль высотного разреза) на баллы главного компонента (PC), суммирующие анатомические вариации древесины, были оценены с использованием подхода множественной регрессии с помощью функции lm in R. Первые три показателя ПК, рассчитанные для набора данных 2012 года (когда были доступны данные о высоте деревьев), использовались в качестве зависимых переменных в множественной регрессии.Для каждого ПК функция Anova в пакете R car (Fox and Weisberg, 2011) использовалась для выполнения обратного выбора модели. Переменная была опущена, когда тест отношения правдоподобия для вложенных моделей показал, что более простая структура модели является более экономной. Кроме того, были проведены тесты Mantel между матрицей попарных географических расстояний и матрицей индивидуальных различий в баллах ПК, чтобы проверить, определяет ли пространственная близость более похожие анатомические черты древесины.Такой анализ был выполнен на уровне графика с использованием пакета R vegan . Влияние климатических колебаний было минимизировано за счет ограничения всех анализов одними и теми же пятью календарными годами (2008–2012 гг.).
Оценка межгодовой стабильности анатомических изменений древесины
PCA был выполнен для каждого набора данных годичных колец анатомических признаков древесины относительно каждого года выбранного периода времени. Чтобы количественно оценить корреляцию индивидуальных анатомических характеристик древесины по годам, был проведен тест Прокруста на показателях ПК за разные годы с использованием функции протест пакета R веганский (Оксанен и др., 2018). Этот статистический тест измеряет корреляцию между двумя конфигурациями точек (то есть баллами ПК). Парные корреляции между всеми возможными комбинациями лет были визуализированы с помощью функции procrustes , показывающей амплитуду и направление изменений оценок PC. Кроме того, для каждого анатомического признака древесины на индивидуальном уровне был рассчитан коэффициент вариации (CV) по годам. Затем индивидуальные CV были усреднены среди людей для ранжирования анатомических характеристик древесины с точки зрения их стабильности во времени.
Результаты
Межиндивидуальные вариации анатомических признаков древесины и их взаимосвязи: одномерный анализ
Ширина годичных колец положительно коррелировала с количеством трахеид в радиальном ряду ( r = 0,99). TRW зависит от ширины EW ( R 2 = 0,993, P <0,001), показывая в среднем процент EW выше 80% (рисунок 2A). TRW также был положительно связан с соотношением радиального и тангенциального диаметров (Drad / Dtan) просвета трахеиды ( R 2 = 0.435, P <0,001) (Рисунок 2B). Размер просвета сохранял пропорциональное увеличение радиального и тангенциального диаметра только в трахеидах EW, но не в трахеидах LW (дополнительный рисунок S3).
Рис. 2. (A) Ширина годичных колец (TRW) пяти крайних годичных колец как функция от процента ранней древесины (EW%). (B) TRW как функция соотношения между радиальным диаметром (Drad) и тангенциальным диаметром (Dtan) просвета трахеиды. Значения отношения Drad / Dtan> 1 указывают на то, что форма трахеиды больше похожа на EW.На вставке представлен графический эскиз трахеид ранней и поздней древесины, как видно в поперечном сечении, с описанием радиального и тангенциального диаметров и их соотношения. Пунктирная линия представляет отношение Drad / Dtan, равное единице. Три символа (круг, ромб и треугольник) соответствуют деревьям из графиков CAM-H (красный), CAM-E (синий) и CAM-L (зеленый). Обе зависимости лучше всего описывались экспоненциальной функцией (*** P <0,001).
Поскольку измерения высоты деревьев до года отбора проб не проводились, только Drad и Dtan внешнего кольца (2012) могут быть связаны с высотой дерева.Dtan был больше связан с высотой дерева, чем Drad, и эти отношения были наиболее сильными для EW (EW Dtan: R 2 = 0,536, EW Drad: R 2 = 0,390 и LW Dtan: R 2 = 0,357, P <0,001) (рисунок 3A). Drad / Dtan масштабируется по высоте дерева: более высокие деревья имели пропорционально больший Dtan в EW ( R 2 = 0,530, P <0,001), что означает, что тангенциальный диаметр увеличивался больше с высотой дерева, чем Drad (Рисунок 3B) .Как следствие, тангенциальное количество трахеид было отрицательно связано с высотой дерева ( R 2 = 0,473, P <0,001).
Рисунок 3. (A) Радиальный диаметр (Drad) просвета трахеиды EW яркого цвета и тангенциальный диаметр (Dtan) просвета трахеиды EW бледного цвета в зависимости от высоты дерева. (B) Отрицательная зависимость между соотношением радиального и тангенциального диаметров просвета трахеиды (Drad / Dtan) и высотой дерева в ярком цвете для части EW и бледном цвете для части LW.Приведены данные за последний год (2012 г.). Каждый символ (круг, ромб и треугольник) соответствует дереву из трех графиков CAM-H (красный), CAM-E (синий) и CAM-L (зеленый). Все взаимосвязи значимы (* P <0,05, ** P <0,01, *** P <0,001).
Диаметр трахеиды влияет как на плотность клеток (CD), количество трахеид на мм 2 , так и на их площадь просвета (LA). И CD, и LA были связаны с высотой дерева, но в противоположных направлениях: CD уменьшается с высотой дерева, а LA увеличивается (дополнительный рисунок S4).
Средний гидравлический диаметр (Dh) и удельная гидравлическая проводимость (Ks) положительно связаны с высотой дерева (дополнительный рисунок S5), тогда как теоретическая гидравлическая проводимость (Kh) – нет. Хотя Kh положительно связан с Drad ( R 2 = 0,503), общее количество трахеид (CNO) ( R 2 = 0,556), EW% ( R 2 = 0,615), базальное приращение площади (BAI) ( R 2 = 0,672), TRW ( R 2 = 0.707) и ширины EW ( R 2 = 0,722) (все взаимосвязи значимы, P <0,001) (Рисунки 4A, B). Кроме того, высокий процент EW соответствовал более высоким значениям индекса «эффективности использования гидравлического углерода» (HCUE) ( R 2 = 0,600, P <0,001) (дополнительный рисунок S6). HCUE линейно увеличивался с TRW ( R 2 = 0,772, P <0,001) и экспоненциально снижался с возрастом ( R 2 = 0.358, P <0,001), что означает, что молодые деревья имеют высокие вложения углерода на единицу проводимости воды.
Рис. 4. (A) Общая проводимость (Kh) как функция ширины ранней древесины (EWW). На вставленных рисунках показаны одинаковые отношения для каждого графика: CAM-H (вверху), CAM-E (в центре) и CAM-L (внизу). Была подобрана линейная функция, за исключением CAM-H, где логарифмическая функция показывает максимальное значение R 2 . (B) Общая проводимость (Kh) как функция процентного содержания ранней древесины (EW%).На вставленных рисунках показаны одинаковые отношения для каждого графика: CAM-H (вверху), CAM-E (в центре) и CAM-L (внизу). Была подобрана экспоненциальная функция. Каждый символ (круг, ромб и треугольник) соответствует дереву из трех графиков CAM-H (красный), CAM-E (синий) и CAM-L (зеленый). Все отношения значимы (*** P <0,001).
Процент Эрливуда (LW%) оказал отрицательное (положительное) влияние на индекс усиления клеточной стенки, измеренный в трахеидах EW ([t / b_ 05 ] 2 ) (Рисунок 5A).Деревья с более плотной древесиной (из-за большего LW%, R 2 = 0,638, P <0,001 - дополнительный рисунок S7) показали более высокие значения [t / b] 2 в части EW (рисунок 5B) . Относительная плотность древесины (RWD) также была отрицательно связана с Drad и Dtan ( R 2 = 0,618 и R 2 = 0,316, соответственно, P <0,001) (дополнительный рисунок S7). как положительно связано с CWT ( R 2 = 0.526, P <0,001).
Рисунок 5. (A) Индекс армирования клеточной стенки ([t / b _ 05 ] 2 ) на участке EW (5-й процентиль) как функция от процента ранней древесины (EW%). Была подобрана логарифмическая функция. (B) Индекс армирования клеточной стенки ([t / b] 2 ) в EW-части (5-й процентиль) как функция относительной плотности древесины в ранней древесине. Каждый символ (круг, ромб и треугольник) соответствует дереву из трех графиков CAM-H (красный), CAM-E (синий) и CAM-L (зеленый).Была подобрана экспоненциальная функция. Все отношения значимы (*** P <0,001).
Толщина клеточной стенки в радиальных клеточных стенках положительно коррелировала с CWT в тангенциальных клеточных стенках (рис. 6). В LW части кольца это соотношение показало более высокий пересечение, в отличие от отношения 1: 1 в EW, с увеличением в тангенциальном направлении сильнее, чем в радиальном направлении. Также была обнаружена слабая, но положительная связь между CWT и возрастом дерева ( R 2 = 0.281, P <0,001), тогда как никакой связи между высотой дерева и CWT не наблюдалось.
Рисунок 6. Толщина стенки ячейки в радиальном направлении (CWT рад) как функция толщины стенки ячейки в тангенциальном (CWT tan) направлении. Фигуры на вставке представляют такое же соотношение для частей EW и LW. Каждый символ (круг, ромб и треугольник) соответствует деревьям с трех графиков CAM-H (красный), CAM-E (синий) и CAM-L (зеленый). Линейные регрессии значительны (*** P <0.001). Пунктирная красная линия представляет соотношение 1: 1.
Межиндивидуальные вариации анатомических признаков древесины и их взаимосвязи: многомерный анализ
PCA по анатомическим признакам древесины относительно годичного кольца 2012 года показало, что первые три PC объясняют 36,8, 28,5 и 13,3% вариации, соответственно (рис. 7). Гидравлические характеристики Kh и HCUE положительно коррелировали с PC1, в то время как [t / b] 2 и [t / b _ 05 ] 2 отрицательно (рисунок 7, дополнительный рисунок S8).Остальные гидравлические характеристики (Dh и Ks) положительно коррелировали с PC2, что означает, что они ортогональны по отношению к другим гидравлическим характеристикам. Все механические характеристики и признаки выделения углерода (таблица 2, дополнительный рисунок S8) показали отрицательную корреляцию с PC1, тогда как PC2 сильно коррелировал с некоторыми анатомическими признаками древесины, связанными с тангенциальным диаметром как в EW, так и LW, а также LA части EW ( Рисунок 7, дополнительный рисунок S8).
Рисунок 7. Результаты PCA по всем анатомическим признакам годичного кольца 2012 года. Стрелки представляют собственное значение каждой переменной для первых двух главных компонентов (PC). Цвета стрелок указывают на функцию, присвоенную каждому анатомическому признаку в соответствии с литературными данными (см. Таблицу 2). Имена стрелок написаны на дополнительном рисунке S8. Три символа (круг, ромб и треугольник) соответствуют индивидуальным оценкам на PC1 ( x -ось) и PC2 ( y -ось) на графиках CAM-H, CAM-E и CAM-L соответственно.
Результаты множественного регрессионного анализа между первыми тремя ПК, а также высота, возраст и площадь деревьев представлены в дополнительной таблице S1. Показатели PC1 увеличивались с высотой дерева (ß = 0,470, P = 0,031) и уменьшались с возрастом (ß = -0,082, P = 0,006; дополнительный рисунок S9A). Показатели PC2 увеличивались с увеличением высоты дерева (ß = 0,546, P <0,001; дополнительный рисунок S9B). Показатели PC3 увеличивались с увеличением высоты дерева (ß = 0,218, P = 0.037) и были достоверно определены графиком ( P = 0,006; дополнительный рисунок S9C). Показатели PC3 значительно различались между CAM-H и CAM-L (прил. P = 0,023). Попарные географические расстояния и попарные различия, рассчитанные на основе оценок PC, были положительно коррелированы только в CAM-H ( r = 0,357, P = 0,019) (дополнительный рисунок S10). Это означает, что в CAM-H деревья, которые расположены пространственно ближе друг к другу, имеют схожие анатомические черты древесины.
Межгодовые изменения анатомических признаков древесины
Анализ главных компонентов был проведен для набора данных относительно каждого кольца дерева (с 2008 по 2012 год), чтобы проверить стабильность анатомических признаков древесины во времени. Тесты Прокруста неизменно показывают, что индивидуальные оценки ПК сильно коррелировали из года в год ( r ≥ 0,8, P = 0,001; рис. 8A). Только 22 из 65 анатомических признаков показали среднее значение CV более 10% и диапазон индивидуальных CV более 20% (Рисунок 8B).Наибольшие межгодовые и межиндивидуальные вариации были показаны [t / b] 2 и LA 1-го и 5-го процентилей (часть LW) (таблица 2, рисунок 8B).
Рисунок 8. (A) Результаты теста Procrustes, анализирующего стабильность результатов ПК во времени. Коэффициенты корреляции и их статистическая значимость показаны на панелях над диагональю. Синие линии представляют собой сдвиги в положении отдельных деревьев по годам. (B) Среднее значение и диапазон индивидуального коэффициента вариации (CV) по годам для каждого анатомического признака.Были отмечены черты, для которых характерны большие различия по годам.
Обсуждение
Внутри древесного кольца: онтогенетические тенденции и анатомия трахеид
Помимо подтверждения того, что более высокие деревья имеют более крупные трахеиды в нижней части ствола, наши результаты продвигают обсуждение вперед. Трахеиды EW у высоких деревьев увеличиваются пропорционально как в радиальном, так и в тангенциальном направлениях. Однако тангенциальный диаметр трахеид частично ограничен диаметром камбиальных материнских клеток и соседними трахеидами.Несмотря на предыдущие работы, подтверждающие, что размер трахеиды увеличивается в первую очередь в радиальном направлении, тогда как тангенциальный диаметр и длина остаются почти постоянными (Skene, 1972; Cuny et al., 2014), здесь мы предполагаем, что радиальный диаметр трахеиды, а тем более их тангенциальный диаметр, в первую очередь зависит от высоты дерева и, в меньшей степени, от высотного положения вдоль разреза. Это подтверждает недавние результаты по хвойным деревьям, растущим на линии деревьев: как только LA трахеид EW стандартизирован для высоты дерева, этот признак не связан с температурой (Kašpar et al., 2019).
Анатомические вариации древесины: функциональные ограничения и компромиссы
Все 65 анатомических признаков древесины в этом исследовании сгруппированы в соответствии с их характеристиками древесины и функцией роста годичных колец, роста клеток, гидравлической эффективности и безопасности, а также механической поддержки и вложения углерода. PCA представляет собой эффективный анализ для обобщения информации, заключенной в большом наборе анатомических характеристик древесины, и показывает высокую степень вариации как между участками, так и внутри них (дополнительный рисунок S11).Мы выделяем две основные анатомические «стратегии» древесины, связанные с первым и вторым ПК, соответственно (рис. 6). С одной стороны, высокие баллы PC1 описывают деревья с высоким процентом EW и гидравлической проводимостью, тогда как низкие баллы PC1 описывают деревья с высоким LW% и высоким RWD, более толстыми стенками трахеид и высоким индексом усиления клеточной стенки, чертами, связанными с гидравлической безопасностью. С другой стороны, PC2 описывает, вкладывается ли каждое дерево в число трахеид или в диаметр их просвета. Высокие баллы PC2 позволяют идентифицировать деревья с небольшим количеством больших трахеид, а низкие баллы PC2 позволяют идентифицировать деревья с большим количеством мелких.Поскольку ПК ортогональны, эта вторая анатомическая стратегия не зависит от первой. PC1 отрицательно связан с возрастом дерева, показывая, что более старые деревья, как правило, имеют более высокую долю LW, более толстую клеточную стенку как в радиальном, так и в тангенциальном направлениях и более высокие LW RWD, чем у молодых деревьев, что может ограничивать их способность к переносу воды, увеличивая инвестиции в углерода и повышают их устойчивость к эмболии (Domec and Gartner, 2002; Rosner et al., 2014), как объясняется в следующем разделе.PC2 сильно, положительно связан с высотой дерева, а это означает, что более высокие деревья, как правило, имеют меньше трахеидов, но имеют больший размер, чтобы максимизировать проводимость воды на единицу площади. Высокие деревья имеют высокий гидравлический диаметр трахеиды и Ks на уровне груди (Rosner et al., 2008; Anfodillo et al., 2012; Olson et al., 2014), тогда как Kh в основном связано с количеством EW: чем выше чем выше процент EW, тем выше проводимость.
Недавние исследования показали чувствительность растений к засухе в зависимости от размера (Bennett et al., 2015; Фахардо и др., 2018; Stovall et al., 2019). Bennett et al. (2015) обнаружили, что засуха более пагубно влияет на рост диаметра ствола больших деревьев. Stovall et al. (2019) сообщили, что высота дерева является наиболее важным предиктором гибели деревьев во время эпизодов засухи, при этом высокие деревья (> 30 м) умирают более чем в два раза, чем короткие деревья (<15 м). Однако высокие или большие деревья не всегда являются старыми деревьями, и отсутствие такой корреляции зависит от характеристик вида (например,g., теневыносливые и светоустойчивые виды) и внутрипопуляционной структуре, такой как местные условия участка, история нарушений дерева, форма роста, а также индивидуальный генотип (Lindenmayer and Laurance, 2016). Наконец, условия участка могут играть важную роль в изменчивости анатомических признаков древесины. Значительный сюжетный эффект обнаружен на PC3; в частности, статистически значимое различие между оценками PC3 CAM-H и CAM-L. Эта разница может свидетельствовать о высотном эффекте, поскольку эти два графика являются наиболее контрастными по высоте (1730 против 1475 м).Тем не менее, PC3 объяснил только 13,3% общих анатомических вариаций древесины между отдельными особями, предполагая, что окружающая среда в любом случае менее важна, чем высота и возраст дерева, на изменение анатомических признаков древесины. Это может быть частично связано с ограниченной протяженностью исследованного высотного градиента (~ 300 м), который, однако, охватывает градиент высоты ели в исследуемой области. По этой причине, возможно, стоит изучить межиндивидуальные вариации анатомических черт древесины на более крупных и воспроизводимых высотных разрезах и в более ограниченных условиях окружающей среды.Слабый, но статистически значимый пространственный эффект обнаружен в CAM-H, где соседние деревья демонстрируют более похожие анатомические черты. В этом случае пространственная близость рассматривается как показатель неоднородности микросреды. Avanzi et al. (2019) продемонстрировали актуальность неоднородности моделей роста микро-окружающей среды на более крупном наборе дендрохронологических данных о деревьях, отобранных на тех же участках. В этом исследовании мы проанализировали 10 деревьев на участке, это не большое количество для полного отражения неоднородности микросреды.
Анатомические признаки древесины на межличностном и межгодовом уровне
Наш анализ межгодовой изменчивости анатомических признаков древесины исследует, какие признаки, скорее всего, не изменятся с течением времени или изменятся незначительно. Результаты PCA сильно коррелированы по годам, что означает, что в целом деревья имеют схожие анатомические черты древесины в их последних сформированных пяти кольцах. Этот результат подчеркивает, в ограниченном временном окне, как деревья демонстрируют глобально согласованные анатомические структуры древесины, даже при различных средних температурах и общем количестве осадков за вегетационный период (дополнительный рисунок S12).Тем не менее, коэффициенты вариации для каждого отдельного анатомического признака древесины показывают, что величина такой межгодовой вариации меняется между признаками и индивидуумами. Мы обнаружили, что индекс армирования клеточной стенки всего кольца, LA LW и LW% являются наиболее вариабельными признаками как на меж-, так и на внутрииндивидуальном уровне. LW, вероятно, является анатомическим признаком древесины, в основном участвующим в потенциально адаптивных реакциях на климат и изменения окружающей среды. Согласно Ларсону (1969), на соотношение EW и LW влияет размер кроны дерева, который, в свою очередь, зависит от плотности древостоя и конкуренции.Когда условия выращивания идеальны, рост в высоту увеличивается, а LW% снижается (Domec and Gartner, 2002). На нашем участке исследования процент EW составляет в среднем> 80%, что указывает на слабое влияние климата на динамику роста отдельных деревьев, как показывает корреляционный анализ климата и роста, выполненный на более крупном наборе данных деревьев, взятых с тех же участков ( Avanzi et al., 2019).
Плотность древесины и гидравлическая уязвимость
Процент поздней древесины влияет на RWD, а также на гидравлические свойства хвойной древесины (Hacke et al., 2001; Домек и Гартнер, 2002 г.). У ели обыкновенной трахеиды EW являются наиболее эффективными водопроводящими трубами с большим количеством ямок на единицу длины канала по сравнению с трахеидами LW (Rosner, 2013). Плотность древесины демонстрирует умеренные или высокие значения наследуемости хвойных пород (например, Hannrup et al., 2004; Martinez Meier et al., 2008; Chen et al., 2018), и, несмотря на то, что эта характеристика не связана напрямую с гидравлической уязвимостью, он участвует в механизмах засухоустойчивости и гидравлических повреждений, при этом более высокая плотность древесины связана с меньшей уязвимостью к кавитации (например,г., Далла-Салда и др., 2009; Роснер, 2013; Рознер и др., 2016; Fernández et al., 2019). Как и в предыдущих исследованиях (Bouche et al., 2014; Bouriaud et al., 2015), настоящие результаты показывают, что RWD в LW больше связан с возрастом дерева, чем с высотой дерева, и что деревья с более высоким RWD в LW имеют более высокий индекс армирования клеточной стенки. Мы использовали индекс усиления клеточной стенки в качестве показателя гидравлической уязвимости (например, Hacke et al., 2001; Rosner, 2013), сделав вывод, что деревья с высоким RWD более устойчивы к трахеидной эмболии.Исследования трахеид LW, EW и переходной древесины предполагают, что, когда имеет место эпизод дефицита воды, сначала возникает кавитация, которая расширяется в трахеидах LW. Только при дальнейшем увеличении дефицита воды эмболия распространяется в EW и, наконец, достигает переходной древесины (Dalla-Salda et al., 2014). Однако потеря проводимости связана не с местом возникновения начальной кавитации, а с анатомической структурой древесины (например, отношением RWD и EW к LW), которая определяет уязвимость видов к засухе (Luss et al., 2019).
Значение для экологических и биологических исследований
В заключение, наши результаты, кажется, последовательно демонстрируют, что в любом исследовании анатомии древесины, экологии и эволюционной генетике крайне важно регистрировать и учитывать высоту дерева для правильного исследования межличностных анатомических вариаций древесины. Таким образом, анатомические характеристики древесины можно стандартизировать на основе высоты дерева, а не диаметра дерева, который не всегда напрямую связан с возрастом дерева. Кроме того, мы также считаем важным учитывать возрастные тенденции, которые могут повлиять на анатомические характеристики древесины с помощью механических поддерживающих функций, таких как CWT, RWD и диаметр просвета трахеид LW.Такие особенности могут в конечном итоге повлиять на устойчивость дерева к эмболии. Мы рекомендуем, чтобы анализ анатомических черт древесины в пределах нескольких годичных колец, но у большого числа людей был хорошим методом для выявления большей части различий как в анатомических характеристиках древесины, так и в условиях микросреды. Эта стратегия позволяет избежать «шума», создаваемого разницей в высоте и возрасте деревьев. Наконец, создание общего сада было бы чрезвычайно полезно для определения того, влияют ли и как отдельные генотипы на анатомические вариации древесины, сводя к минимуму неоднородность микросайтов.
Заявление о доступности данныхНаборы данных, созданные для этого исследования, доступны по запросу соответствующему автору.
Взносы авторов
Все авторы спланировали и разработали исследование. AlP, AnP, CA и CU провели полевые исследования. Данные были собраны AlP. AlP, GA, AnP, CA и AC проанализировали данные. AlP руководил написанием рукописей при участии всех авторов. Все авторы высказались критически о черновиках и дали окончательное одобрение для публикации.
Финансирование
AlP была поддержана Политехническим университетом Марке (Италия) для ее постдока за рубежом (2016 год) в Швейцарском федеральном исследовательском институте (WSL), который обеспечил техническую и научную поддержку. PF и GA были поддержаны швейцарским вкладом Государственного секретариата по образованию, исследованиям и инновациям (SERI) в проект ЕС h3020 GenTree (№ 676876). UB получил финансирование от проекта «SustES – стратегии адаптации для устойчивых экосистемных услуг и продовольственной безопасности в неблагоприятных экологических условиях» (Чехия.02.1.01 / 0.0 / 0.0 / 16_019 / 0000797). AnP и CA были поддержаны проектом ERAnet BiodivERsA «TipTree» (ANR-12-EBID-0003).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Мы хотели бы поблагодарить Paul J. Krusic за его комментарии, Ufficio per la Biodiversità в Пистойе и Итальянскую государственную лесную службу Abetone (PT) за логистическую поддержку полевых работ.Благодарим Валерию Галлуччи, Маттео Гарбарино, Кристину Леонардузи и Даниэле Кастагнери за помощь во время полевых исследований, а также Сандро Морганти, Марию Селесте Лабриола, Катию Богги и Иларию Спану за техническую помощь во время лабораторной работы.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fpls.2020.00683/full#supplementary-material
Сноски
Список литературы
Алони Р.(2013). «Роль гормонов в контроле дифференцировки сосудов», в Клеточные аспекты образования древесины , изд. Дж. Фромм, (Берлин: Springer), 99–139. DOI: 10.1007 / s00425-013-1927-8
CrossRef Полный текст | Google Scholar
Анфодилло Т., Деслорье А., Менарди Р., Тедольди Л., Пети Г. и Росси С. (2012). Расширение каналов ксилемы у хвойного дерева зависит от более длительного времени распространения клеток вниз по стволу. J. Exp. Бот. 63, 837–845.DOI: 10.1093 / jxb / err309
CrossRef Полный текст | Google Scholar
Анфодилло Т., Пети Г. и Кривелларо А. (2013). Расширение осевого канала у древесных пород: анатомическая структура, которой еще не уделяется должного внимания. IAWA J. 34, 352–364.
Google Scholar
Avanzi, C., Piermattei, A., Piotti, A., Büntgen, U., Heer, K., Opgenoorth, L., et al. (2019). Определение влияния пространственной близости и генетического сходства на индивидуальные показатели роста в природных популяциях ели европейской. Sci. Total Environ. 650, 493–504. DOI: 10.1016 / j.scitotenv.2018.08.348
CrossRef Полный текст | Google Scholar
Бекман, Х. (2016). Анатомия древесины и экология по признакам. IAWA J. 37, 127–151.
Google Scholar
Беннетт А.С., МакДауэлл Н.Г., Аллен К.Д. и Андерсон-Тейшейра К.Дж. (2015). Крупные деревья больше всего страдают от засухи в лесах во всем мире. Nat. Растения 1: 15139. DOI: 10.1038 / nplants.2015.139
CrossRef Полный текст | Google Scholar
Буш, П.S., Larter, M., Domec, J.C., Burlett, R., Gasson, P., Jansen, S., et al. (2014). Общий обзор гидравлической и механической безопасности ксилемы хвойных пород. J. Exp. Бот. 65, 4419–4431. DOI: 10.1093 / jxb / eru218
CrossRef Полный текст | Google Scholar
Бурио, О., Теодосиу, М., Кирдьянов, А. В., Вирт, К. (2015). Влияние плотности древесины на оценки годовой урожайности на основе годичных колец и ее ошибки в ели европейской. Биогеонауки 12, 6205–6217.
Google Scholar
Каррер М., Брунетти М. и Кастагнери Д. (2016). Отпечаток экстремальных климатических явлений в вековых временных рядах анатомических особенностей древесины высокогорных хвойных пород. Фронт. Plant Sci. 7: 683. DOI: 10.3389 / fpls.2016.00683
CrossRef Полный текст | Google Scholar
Каррер М., фон Аркс Г., Кастагнери Д. и Пети Г. (2015). Извлечение аллометрической и экологической информации из временных рядов размера канала: проблема стандартизации и ее связь с древовидной гидравлической архитектурой. Tree Physiol. 35, 27–33. DOI: 10.1093 / treephys / tpu108
CrossRef Полный текст | Google Scholar
Кастагнери Д., Фонти П., фон Аркс Г. и Каррер М. (2017). Как климат влияет на морфогенез ксилемы в течение вегетационного периода? Выводы из долгосрочной внутрикольцевой анатомии у Picea abies. Ann. Бот. 119, 1011–1020. DOI: 10.1093 / aob / mcw274
CrossRef Полный текст | Google Scholar
Чен, З. К., Байсон, Дж., Пан, Дж., Карлссон, Б., Андерссон, Б., Вестин, Дж. И др. (2018). Точность геномной селекции по признакам роста и качества древесины в двух испытаниях потомства с контрольным опылением с использованием захвата экзома в качестве платформы для генотипирования ели европейской. BMC Genomics 19: 946. DOI: 10.1186 / s12864-018-5256-y
CrossRef Полный текст | Google Scholar
Кьяруги А. (1936). L’indigenato della «Picea excelsa» nell’Appennino Etrusco. Nuovo G. Bot. Ital. 63, 131–166.
Google Scholar
Куни, Х.Э., Ратгебер, К. Б., Франк, Д., Фонти, П., и Фурнье, М. (2014). Кинетика развития трахеид объясняет кольцевую структуру хвойных деревьев. New Phytol. 203, 1231–1241. DOI: 10.1111 / Nph.12871
CrossRef Полный текст | Google Scholar
Далла-Салда, Г., Фернандес, М. Э., Сержент, А. С., Розенберг, П., Бадель, Э. и Мартинес Мейер, А. (2014). Динамика кавитации в кольце дерева Дугласа-ели: переходное дерево, властелин кольца? J. Plant Hydr. 1: e0005.
Google Scholar
Далла-Салда, Г., Мартинес-Мейер, А., Кочард, Х., и Розенберг, П. (2009). Изменение плотности древесины и гидравлических свойств клонов пихты дугласской ( Pseudotsuga menziesii (Mirb.) Franco), связанных с волной жары и засухи во Франции. Для. Ecol. Manag. 257, 182–189.
Google Scholar
Де Микко В., Каррер М., Ратгебер К. Б. К., Камареро Дж. Дж., Вольтас Дж., Керубини П. и др. (2019). От ксилогенеза до годичных колец: характеристики древесины для изучения реакции дерева на изменения окружающей среды. IAWA J. 40, 155–182.
Google Scholar
Денн, М. П. (1988). Определение поздней древесины по Морку (1928). IAWA Bull. 10, 59–62.
Google Scholar
Домек, Дж. К., и Гартнер, Б. Л. (2002). Чем отличается водный транспорт и хранение воды у ранней и поздней древесины хвойных пород? J. Exp. Бот. 53, 2369–2379. DOI: 10.1093 / jxb / erf100
CrossRef Полный текст | Google Scholar
Domec, J. C., Уоррен, Дж. М., Мейнзер, Ф. К., и Лахенбрух, Б. (2009). Защита от разрушения ксилемы за счет взрыва и посева воздуха в корнях, стволах и ветвях молодых и старых хвойных деревьев. IAWA J. 30, 101–120.
Google Scholar
Энквист, Б. Дж. (2002). Универсальное масштабирование в аллометрии деревьев и сосудистых растений: к общей количественной теории, связывающей форму и функцию растений от клеток до экосистем. Tree Physiol. 22, 1045–1064. DOI: 10,1093 / treephys / 22.15-16.1045
CrossRef Полный текст | Google Scholar
Эванс, М. Е. К., Гуггер, П. Ф., Линч, А. М., Гитерман, К. Х., Фаулер, Дж. К., Клесс, С. и др. (2018). Дендроэкология встречается с геномикой в общем саду: новые взгляды на адаптацию к климату. New Phytol. 218, 401–403. DOI: 10.1111 / Nph.15094
CrossRef Полный текст | Google Scholar
Эверт, Р. Ф. (2006). Анатомия растений, меристемы, клетки и ткани растительного тела Исава – их структура, функции и развитие. Хобокен: Уайли.
Google Scholar
Фахардо, А., Макинтайр, Э. Дж. Б. и Олсон, М. Э. (2018). Когда невысокий рост является преимуществом на деревьях. Trends Ecol. Evol. 34, 193–199. DOI: 10.1016 / j.tree.2018.10.011
CrossRef Полный текст | Google Scholar
Фернандес, М. Э., Баротто, А. Дж., Мейер, А. М., Дьенге, Дж. Э., Тесон, Н., Киньонес Марторелло, А. С. и др. (2019). Новый взгляд на анатомию древесины и взаимосвязь функций: как эвкалипт бросает вызов тому, что мы уже знаем. Для. Ecol. Manag. 454: 117638.
Google Scholar
Фонти П., Брюханова М. В., Мыглан В. С., Кирдянов А. В., Наумова О. В., Ваганов Е. А. (2013). Температурные реакции структуры ксилемы Larix sibirica (Pinaceae) из Российского Алтая. Am. J. Bot. 100, 1332–1343. DOI: 10.3732 / ajb.1200484
CrossRef Полный текст | Google Scholar
Фонти П., Янсен С. (2012). Пластичность ксилемы в зависимости от климата. New Phytol. 195, 734–736. DOI: 10.1111 / j.1469-8137.2012.04252.x
CrossRef Полный текст | Google Scholar
Фокс, Дж., И Вайсберг, С. (2011). Приложение R к прикладной регрессии, второе издание. Thousand Oaks, CA: Wiley.
Google Scholar
Gartner, B. L. (1995). «Паттерны изменений ксилемы в дереве и их гидравлические и механические последствия», в Стебли растений: физиология и функциональная морфология , изд.Б. Л. Гартнер, (Сан-Диего: Academic Press), 125–149.
Google Scholar
Gärtner, H., Cherubini, P., Fonti, P., von Arx, G., Schneider, L., Nievergelt, D., et al. (2015a). Техническая перспектива в современных исследованиях древесных колец – как преодолеть дендроэкологические и анатомические проблемы древесины. J. Vis. Exp. 97: e52337. DOI: 10.3791 / 52337
CrossRef Полный текст | Google Scholar
Гертнер Х., Лучинетти С. и Швайнгрубер Ф. Х. (2015b).Новый санный микротом, сочетающий в себе анатомию древесины и экологию годичных колец. IAWA J. 36, 452–459.
Google Scholar
Гертнер Х., Швайнгрубер Ф. Х. (2013). Микроскопические методы подготовки к анализу стеблей растений. Бирменсдорф: Verlag-WSL.
Google Scholar
Хаке, У. Г., Лахенбрух, Б., Питтерман, Дж., Майр, С., Домек, Дж. К., и Шульте, П. Дж. (2015). «Гидравлическая архитектура хвойных пород», в Функциональная и экологическая анатомия ксилемы , изд.У. Г. Хаке (Швейцария: Springer International Publishing), 39–75.
Google Scholar
Хаке, У. Г., Сперри, Дж. С., Покман, У. Т., Дэвис, С. Д., и МакКалло, К. А. (2001). Тенденции изменения плотности и структуры древесины связаны с предотвращением взрыва ксилемы под действием отрицательного давления. Oecologia 126, 457–461. DOI: 10.1007 / s004420100628
CrossRef Полный текст | Google Scholar
Ханнруп, Б., Кахалан, К., Шантр, Г., Грабнер, М., Карлссон, Б., Le Bayon, I., et al. (2004). Генетические параметры роста и признаки качества древесины у Picea abies . Сканд. J. For. Res. 19, 14–29.
Google Scholar
Heer, K., Behringer, D., Piermattei, A., Bässler, C., Brandl, R., Fady, B., et al. (2018). Связывание дендроэкологии и генетики ассоциации в естественных популяциях: ответы на стресс, заархивированные в годичных кольцах, связаны с генотипами SNP у пихты пихты ( Abies alba, Mill.). Мол. Ecol. 27, 1428–1438. DOI: 10.1111 / mec.14538
CrossRef Полный текст | Google Scholar
Jyske, T., и Hölttä, T. (2015). Сравнение гидравлической архитектуры флоэмы и ксилемы в стеблях Picea abies. New Phytol. 205, 102–115. DOI: 10.1111 / nph.12973
CrossRef Полный текст | Google Scholar
Кашпар, Дж., Анфодилло, Т., и Тремл, В. (2019). Размер дерева в основном влияет на изменение характеристик ксилемы в экотоне линии деревьев. Деревья 33, 1657–1665.
Google Scholar
Лахенбрух Б., Мур Дж. Р. и Эванс Р. (2011). «Радиальные вариации в структуре и функциях древесины у древесных растений и гипотезы их возникновения» в «Изменения в структуре и функциях дерева, связанные с размером и возрастом» , ред. FC Meinzer, B. Lachenbruch и TE Dawson, (Dordrecht: Спрингер), 121–164.
Google Scholar
Ларсон П. Р. (1969). Формирование древесины и понятие качества древесины. Нью-Хейвен, Коннектикут: Йельский университет.
Google Scholar
Ларсон П. Р. (1994). Сосудистый камбий, развитие и строение. Берлин: Springer-Verlag.
Google Scholar
Лаццарин М., Кривелларо А., Уильямс К. Б., Доусон Т. Э., Моцци Г. и Анфодилло Т. (2016). Анатомия трахеиды и ямки у высокого дерева Sequoiadendron giganteum различается в тандеме. IAWA J. 37, 172–185.
Google Scholar
Линденмайер, Д. Б., и Лоранс, В.Ф. (2016). Уникальные проблемы сохранения больших старых деревьев. Trends Ecol. Evol. 31, 416–418. DOI: 10.1016 / j.tree.2016.03.003
CrossRef Полный текст | Google Scholar
Luss, S., Lundqvist, S., Evans, R., Grahn, T., Olsson, L., Petit, G., et al. (2019). Изменчивость структуры древесины внутри кольца и ее связь с засухой. IAWA J. 40, 288–310. DOI: 10.1163 / 22941932-401
CrossRef Полный текст | Google Scholar
Магри, Д., Agrillo, E., Di Rita, F., Furlanetto, G., Pini, R., Ravazzi, C., et al. (2015). Голоценовая динамика популяций древесных таксонов в Италии. Rev. Palaeobot. Палинол. 218, 267–284.
Google Scholar
Мальпиги, М. (1675). Anatome Plantarum. Лондон: Мартин.
Google Scholar
Мартинес Мейер, А. Г., Санчес, Л., Салда, Г., Пасторино, М. Дж. М., Готри, Ж.-Й., Галло, Л. А., и др. (2008). Генетический контроль кольцевой реакции пихты Дугласовой ( Pseudotsuga menziesii (Mirb.) Franco) к засухе и аномальной жаре 2003 г. во Франции. Ann. Для. Sci. 65, 102–102.
Google Scholar
МакКулло, К., Сперри, Дж. С., Лахенбрух, Б., Мейнцер, Ф. К., Райх, П. Б. и Фелькер, С. (2010). Скважина с движущейся водой: сравнение гидравлической эффективности веток и стволов хвойных, кольцевидных и диффузно-пористых саженцев умеренных и тропических лесов. New Phytol. 186, 439–450. DOI: 10.1111 / j.1469-8137.2010.03181.x
CrossRef Полный текст | Google Scholar
Оксанен, Я., Бланше, Ф. Г., Френдли, М., Киндт, Р., Лежандр, П., МакГлинн, Д. и др. (2018). Веган: Пакет «Экология сообщества». Версия пакета R 2.4–6.
Google Scholar
Олсон, М. Э., Анфодилло, Т., Роселл, Дж. А., Пети, Г., Кривелларо, А., Иснард, С., и др. (2014). Универсальная гидравлика цветковых растений: диаметр сосуда зависит от длины стебля по родословным, повадкам и климату покрытосеменных. Ecol. Lett. 17, 988–997. DOI: 10.1111 / ele.12302
CrossRef Полный текст | Google Scholar
Пломион, К., Лепровост, Г., и Стокс, А. (2001). Образование древесины на деревьях. Plant Physiol. 127, 1513–1523.
Google Scholar
Прендин А. Л., Пети Г., Каррер М., Фонти П., Бьерклунд Дж. И фон Аркс Г. (2017). Новые перспективы исследований, основанные на новом подходе к количественной оценке толщины стенки трахеиды. Tree Physiol. 37, 976–983. DOI: 10.1093 / treephys / tpx037
CrossRef Полный текст | Google Scholar
Prendin, A. L., Petit, G., Fonti, P., Риксен, К., Даус, М.А., и фон Аркс, Г. (2018). Осевая архитектура ксилемы Larix decidua , подвергшаяся обогащению CO2 и нагреванию почвы на границе дерева. Функц. Ecol. 32, 273–287.
Google Scholar
R Основная команда (2018). R: язык и среда для статистических вычислений . Вена: Фонд R для статистических вычислений.
Google Scholar
Роселл, Дж. А., Олсон, М. Э., и Анфодилло, Т. (2017). Масштабирование диаметра сосуда ксилемы с размером растения: причины, прогнозы и нерешенные вопросы. Curr. Лес. Rep. 3, 46–59.
Google Scholar
Рознер, С. (2013). Гидравлическая и биомеханическая оптимизация ствола древесины европейской ели: обзор. IAWA J. 34, 365–390.
Google Scholar
Рознер С., Кляйн А., Мюллер У. и Карлссон Б. (2008). Компромисс между реакцией на гидравлическую и механическую нагрузку зрелой древесины ствола ели европейской. Tree Physiol. 28, 1179–1188. DOI: 10.1093 / treephys / 28.8.1179
CrossRef Полный текст | Google Scholar
Роснер, С., Svetlik, J., Andreassen, K., Borja, I., Dalsgaard, L., Evans, R., et al. (2014). Плотность древесины как критерий засухоустойчивости ели обыкновенной. Кан. J. For. Res. 44, 154–161.
Google Scholar
Rosner, S., Světlík, J., Andreassen, K., Børja, I., Dalsgaard, L., Evans, R., et al. (2016). Новые модели гидравлической уязвимости для северных хвойных пород показывают, что у оппортунистов могут быть более низкие шансы на выживание в условиях экстремальных климатических явлений. Фронт.Plant Sci. 7: 831. DOI: 10.3389 / fpls.2016.00831
CrossRef Полный текст | Google Scholar
Рунгваттана, К., Хиц, П. (2017). Радиальные вариации функциональных характеристик древесины отражают связанные с размерами адаптации механики и гидравлики дерева. Функц. Ecol. 32, 1–13.
Google Scholar
Скин, Д. С. (1972). Кинетика развития трахеид у Tsuga canadensis Carr и ее связь с энергией дерева. Ann. Бот. 36, 179–187.
Google Scholar
Стовалл, А. Э. Л., Шугарт, Х., Си Ю. (2019). Высота дерева объясняет риск гибели во время сильной засухи. Nat. Commun. 10: 4385. DOI: 10.1038 / s41467-019-12380-6
CrossRef Полный текст | Google Scholar
Тайри, М. Т., и Эверс, Ф. У. (1991). Гидравлическая архитектура деревьев и других древесных растений. New Phytol. 119, 345–360.
Google Scholar
Ваганов Э.А., Хьюз М.К., Шашкин А.В. (2006). «Сезонная активность камбия и производство новых клеток ксилемы», в Динамика роста хвойных колец, экологические исследования (анализ и синтез) , изд. Е. А. Ваганов, (Берлин: Springer), 71–104.
Google Scholar
фон Аркс, Г., и Каррер, М. (2014). ROXAS – новый инструмент для построения многовековой хронологии трахеиды и просвета хвойных пород. Дендрохронология 32, 290–293.
Google Scholar
фон Аркс, Г., Кривелларо, А., Прендин, А. Л., Чуфар, К., и Каррер, М. (2016). Количественная анатомия древесины – практические рекомендации. Фронт. Plant Sci. 7: 781. DOI: 10.3389 / fpls.2016.00781
CrossRef Полный текст | Google Scholar
Вест, Г. Б., Браун, Дж. Х. и Энквист, Б. Дж. (1997). Общая модель происхождения законов аллометрического масштабирования в биологии. Наука 276, 122–126. DOI: 10.1126 / science.276.5309.122
CrossRef Полный текст | Google Scholar
Ель обыкновенная – обзор
Возникновение и структура
Лайхо и Микола (1964) исследовали начало микоризной колонизации у сеянцев Pinus sylvestris и Picea abies .У Picea развилась нормальная эктомикориза, но на Pinus образовались грубая сеть Хартига, внутриклеточная колонизация и тонкая или отсутствующая грибковая мантия. Эти эктендомикоризы изначально были очень многочисленны и неизменно присутствовали на Pinus на грядках из почвы сельскохозяйственного происхождения, но действительно были очень редко на Picea. Самым ранним признаком колонизации было формирование сети Хартига, которая следовала за апикальной меристемой по мере роста корня.Вследствие этого интенсивность внутриклеточного проникновения увеличивалась по направлению к более старой части корня, так что клетки были почти заполнены клубками перегородочных гиф шириной до 15 мкм каждая. Эти эктендомикоризные корни сохранялись не менее года с небольшими признаками дегенерации гиф. Как писал Микола (1965): «Внутриклеточные гифы не повреждают корковые клетки; наблюдали, что как растительные клетки, так и внутриклеточные гифы живут по крайней мере один год после начала колонизации; даже ядра таких сильно колонизированных кортикальных клеток были ясно видны на окрашенных препаратах ».
Scales and Peterson (1991a) провели подробное исследование структуры и развития эктендомикоризы Pinus Banksiana , образованной Wilcoxina mikolae var. миколаэ. Возникающие короткие корни покрываются гифами, которые, кажется, встроены в матричный материал (рис. 7.1a. Гифы в этом матриксе сильно разветвлены (рис. 7.1b. торчащие корневые волоски.На этом этапе только несколько гиф узкого диаметра пересекают верхушку корня (рис.1c. В зрелой структуре (рис. 7.1d, 7.2) верхушка полностью закрыта, за исключением тех случаев, когда боковые корни растут очень быстро. Как показано на рис. 7.2a, сразу за вершиной начинает формироваться однорядная сеть Хартига. Сначала он проникает между эпидермальными и внешними кортикальными клетками, но в конечном итоге распространяется на внутреннюю кору, прилегающую к энтодерме. Сеть Хартига имеет лабиринтную структуру (рис. 7.2b, типичная для эктомикоризы. Одна или две клетки, удаленные от тех, в которых наблюдается самое раннее образование сети Хартига, происходит внутриклеточное проникновение (рис. 7.2а. Гифы, проникнув в клетку, многократно разветвляются (рис. 7.3).
Рисунок 7.1. Сканирующая электронная микроскопия коротких корней Pinus Banksiana-Wilcoxina mikolae var mikolae mycorrhizas. (а) Молодой надводный короткий корень, покрытый гифами (отмечен стрелкой), которые, по-видимому, частично встроены в матричный материал (*). (b) Субапикальная область микоризного короткого корня, демонстрирующая сильно разветвленные гифы (наконечники стрелок), внедренные в матричный материал (двойные наконечники стрелок) на поверхности корня.(c) Короткий корень, показывающий апикальные корневые волоски (большие стрелки) и гифы (наконечники стрелок), контактирующие с поверхностью корня в зоне колонизации. Гифы небольшого диаметра (двойные стрелки) пересекают поверхность корня. (d) Апикальная область моноподиальных микоризных корней, показывающая мантию (*). Полосы = 100 мкм.
От Scales and Peterson (1991a), с разрешения Авторские права © 1991Рисунок 7.2. Световая микроскопия дихотомических коротких корней Pinus banksiana-Wilcoxina mikolae var mikolae mycorrhizas.(а) Продольный разрез, показывающий сеть Хартига (стрелки), оболочку (*) и внутриклеточные гифы (двойные стрелки). Полоса = 100 мкм. (b) Более сильное увеличение области главной оси корня, показанной на (a). Гифы как в сети Хартига (наконечники стрелок), так и внутриклеточных гиф (двойные наконечники стрелок) довольно вакуолизированы. Полоса = 50 мкм.
От Scales and Peterson (1991a), с разрешения Авторские права © 1991Рисунок 7.3. Сканирующая электронная микроскопия одиночной кортикальной клетки микоризы, образованной между Pinusbankiana и Wilcoxina mikolae var mikolae. Видно, что внутриклеточные гифы (наконечники стрелок) разветвлены, а сеть Хартига (черные наконечники стрелок) однотонная. Полоса = 100 мкм.
От Scales and Peterson (1991a), с разрешения. Авторское право © 1991Все аспекты структуры и развития этой эктендомикоризы, за исключением внутриклеточного проникновения, по-видимому, аналогичны тем, которые наблюдаются при эктомикоризе. Когда тот же самый гриб был использован для инокуляции Picea mariana и Betula alleghaniensis (Scales and Peterson, 1991b), были получены эктомикоризы, типичные для грибов, образованных другими грибами.Хорошо развитая сеть Хартига с лабиринтным ростом наблюдалась в обоих типах, но ни одна из них не показала никаких доказательств внутриклеточного проникновения. Эти описания показывают, что утверждение, что кортикальные клетки мертвы и лишены цитоплазмы в корнях EECM, в целом неверно. Ни Микола (1965), ни Скейлс и Петерсон (1991a) не обнаружили никаких признаков «переваривания» гиф.
Микола пришел к выводу, что вид эктендомикоризы, образованной грибами E-штамма (см. Ниже), был характерен для молодых 1-3-летних Pinus в почве питомников в широком диапазоне плодородия почвы, кислотности и содержания гумуса и что интенсивность их образования не сильно зависела от силы света.Таким образом, продолжительность жизни этого вида микоризы сравнялась с продолжительностью жизни типичных эктомикориз без внутриклеточной колонизации (один год и более) и была во много раз дольше, чем у немикоризных корней аналогичного положения в корневой системе.
Лайхо (1965) провел широкое исследование эктендомикоризы в питомниках и лесах Европы и Америки и экспериментально синтезировал микоризу на ряде видов деревьев с использованием выделенных из них грибов Е-штамма. Он столкнулся только с одной формой эктендомикоризы, у которой корни были населены грубым перегородчатым мицелием, который образовывал сильную сеть Хартига и внутриклеточную колонизацию.Он пришел к выводу, что грибы E-штамма присутствовали во всех эктендомикоризах, которые он подробно исследовал, но что должны быть и другие возбудители грибов, так как грибы E-штамма колонизировали только виды Pinus и Larix, , тогда как некоторые эктендомикоризы, несомненно, имели место. по видам Picea и другим родам. Однако он подчеркнул, что существует вероятность ошибки, поскольку стареющие микоризы часто имеют внутриклеточную колонизацию, и он процитировал наблюдение Миколы (1948) о том, что Cenococcum может проникать через стенки кортикальных клеток растений, растущих в неподходящих условиях.Этот момент еще более подтверждается наблюдением, что вторжение в стареющие корковые клетки – обычное дело (Harley, 1936; Atkinson, 1975; Nylund, 1981). Лайхо был впечатлен отсутствием вредного воздействия колонизации EECM на сеянцев Pinus и пришел к выводу, что это был «сбалансированный симбиоз», который уступил место эктомикоризе по мере старения сеянца, особенно в лесных условиях.
Wilcox (1968b, 1971) описал подобные эктендомикоризы на молодых растениях P.resinosa , у которых грубые гифы образуют сеть Хартига как в длинных, так и в коротких корнях. Эти гифы имеют тенденцию закручиваться по спирали в клеточной стенке, следуя углу фибрилл целлюлозы. Они проникают в клеточную стенку через участки ямок, а также посредством довольно сложных аппрессорий и прорастают в просвет клетки. Гифы, по-видимому, стимулируются к межклеточному распространению за вершиной длинных корней и вокруг развивающихся боковых сторон, но не вторгаются в меристемы. Они могут быть активны вокруг боковых инициалов и колонизировать молодые корни, когда они проходят через кору.Хотя эктендомикоризы часто присутствуют на большинстве коротких корней молодых проростков в почвах питомников, они позже заменяются эктомикоризами, образованными тонкими гифами, вероятно, другого гриба.
Повсеместное сокращение роста еловых лесов в результате воздействия ветра
Абстрактные
Фон
В последние десятилетия во многих лесных экосистемах по всему миру значительно возросла частота и сила естественных нарушений, например, сильных ветров и нашествий насекомых.Ожидается, что в будущем изменение климата приведет к дальнейшему усилению режимов возмущений, что делает устранение нарушений в управлении экосистемами главным приоритетом. В качестве предварительного условия необходимо более широкое понимание воздействий нарушений и реакции экосистемы. Что касается воздействия сильных ветров – самого опасного фактора беспокойства в Европе, – мониторинг и управление сосредоточены на структурных повреждениях, то есть гибели деревьев от выкорчевывания и поломки стволов. Влияние на функционирование деревьев, переживших шторм (например,g., их продуктивность и распределение) до настоящего времени редко учитывались.
Методология / основные результаты
Здесь мы показываем, что сокращение прироста было значительным и повсеместным в лесном ландшафте площадью 6,79 млн га на юге Швеции после шторма Гудрун (январь 2005 г.). Связанное с ветром сокращение роста еловых ( Picea abies (L.) Karst.) Лесов, переживших шторм, превысило 10% в наиболее пострадавших регионах и было тесно связано с максимальной скоростью порыва ветра (R 2 = 0.849) и повреждение конструкции ветром (R 2 = 0,782). В ландшафтном масштабе снижение прироста, связанное с ветром, составило 3,0 млн. М 3 за три года после Гудрун. Таким образом, он превышает вторичный ущерб от жуков-короедов после Гудруна, а также долгосрочный средний ущерб от выкорчевывания и поломки стебля в Швеции.
Выводы / Значение
Мы пришли к выводу, что воздействие сильных ветров на лесные экосистемы не ограничивается непосредственно видимой зоной структурного повреждения, и призываем к более широкому рассмотрению возмущающего воздействия на структуру экосистемы и , функционирующую в контексте управления лесами и смягчения последствий изменения климата. .
Образец цитирования: Зейдл Р., Бленнов К. (2012) Повсеместное сокращение роста еловых лесов норвежской в результате воздействия ветра. PLoS ONE 7 (3): e33301. https://doi.org/10.1371/journal.pone.0033301
Редактор: Бен Бонд-Ламберти, Тихоокеанская северо-западная национальная лаборатория Министерства энергетики США, США
Поступила: 25 ноября 2011 г .; Принята к печати: 10 февраля 2012 г .; Опубликовано: 7 марта 2012 г.
Авторские права: © 2012 Seidl, Blennow.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Руперт Зайдл был поддержан стипендией Марии Кюри в рамках Седьмой рамочной программы Европейского сообщества («iLand – основа для индивидуального моделирования лесных ландшафтов в условиях меняющегося климата и возмущений», грант 237085).Исследование было дополнительно поддержано проектом ЕС MOTIVE («Модели адаптивного управления лесами», грант 226544) (www.motive-project.net) и фондом программы стратегических исследований окружающей среды Mistra-SWECIA (www.mistra-swecia.se ). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
В последние десятилетия в лесах по всему миру участились природные нарушения и их масштабы [1], [2].Как следствие, нарушения, например лесные пожары, нашествия насекомых и сильные ветры, становятся все более серьезной проблемой для устойчивого управления лесными экосистемами. Нарушения могут иметь сильные негативные последствия для производства древесины и экономики, основанной на древесине, например, из-за девальвации древесины, необходимости преждевременного сбора урожая и перенасыщения рынка бобовыми. Единичный шторм 8 января -го –9 -го 2005 года (шторм «Гудрун»), например, по оценкам, нанес общий экономический ущерб в 2 раза.4 миллиарда евро только в лесном хозяйстве Швеции [3]. Кроме того, возмущающие явления могут превратить леса, являющиеся стоком углерода (C) в атмосферу, в источник углерода [4], [5]. Таким образом, они могут серьезно помешать достижению целей смягчения последствий изменения климата посредством управления лесным хозяйством. В свою очередь, антропогенное изменение климата также влияет на режимы возмущений. Наблюдаемое усиление возмущений частично связано с недавними изменениями в климатической системе [6], [7], а прогнозы будущих климатических сценариев указывают на дальнейшее увеличение возмущений [8], [9].Таким образом, устранение нарушений становится все более центральной проблемой в науках об устойчивости в целом и в управлении экосистемами в частности [10].
Сильные ветры – самый важный фактор нарушения лесных экосистем Европы (судя по объему поврежденной древесины, см. [1]). Воздействие ветра на деревья разнообразно и варьируется от их выкорчевывания с корнем до механического повреждения отдельных участков деревьев (рис. 1). Воздействие ветра на структуру и демографию лесов, т. Е., смертность в результате выкорчевывания и поломки стеблей до настоящего времени была главной проблемой в контексте управления лесным хозяйством и экосистемами. Они хорошо видны на ландшафте и имеют значительную величину, составляя в среднем 18,7 млн. М. [1]. Однако и на уцелевшие деревья и насаждения в ландшафтах, подверженных сильным ветрам, также действует физическая сила таких экстремальных атмосферных явлений.В экологической литературе, посвященной их влиянию на остаточную популяцию деревьев, сообщается как об увеличении (из-за выброса убитых ветром соседей и последующем увеличении доступности ресурсов для выживших деревьев, см., Например, [11] – [13]), так и об уменьшении (из-за механического повреждения ветвей, корней и ткани ксилемы, см., например, [14], [15]) в росте и продуктивности в ответ на сильный ветер. Кроме того, было обнаружено, что механическое воздействие ветра значительно изменяет распределение углеводов в деревьях, увеличивая выделение в подземные компартменты [16], [17].Однако эти воздействия на фундаментальные экофизиологические процессы в выжившей популяции деревьев, обобщенные здесь как функциональные воздействия, на сегодняшний день не учитываются при оценке ущерба от ветра в контексте лесного хозяйства и устойчивого лесопользования, в основном потому, что один шторм может иметь одновременно положительные и отрицательные эффекты на уровне отдельных деревьев [18], а общий знак и величина функционального воздействия на лесные ландшафты до сих пор не решены. Тем не менее, игнорирование таких функциональных эффектов может привести к значительно предвзятой оценке ветровых возмущений, например.g., что касается воздействия на производство древесины и хранение углерода, в управлении экосистемами.
Рис. 1. Воздействие сильных ветров и их влияние на структуру и функционирование лесных экосистем.
Штормы могут оказывать различное воздействие на деревья – от выкорчевывания с корнем до механического повреждения различных участков деревьев. Эти воздействия приводят к двум различным типам реакций: влиянию на структуру экосистемы (например, через гибель деревьев) и функциональной реакции (т.е. изменениям в фундаментальных экофизиологических процессах).Структурные эффекты связаны с изменениями в демографии и составе, в то время как функциональные эффекты касаются реакции существующей / оставшейся растительности и почвы. Оценки ветровых воздействий в контексте управления лесным хозяйством и экосистемами на сегодняшний день в основном сосредоточены на первом, в то время как здесь мы оцениваем значимость и величину функционального воздействия сильных ветров в масштабе ландшафта, уделяя особое внимание краткосрочным изменениям роста деревьев. Обратите внимание, что эта схематическая иллюстрация фокусируется только на эффектах первого порядка; структурные эффекты будут влиять на функционирование (и наоборот) в ходе динамики леса.
https://doi.org/10.1371/journal.pone.0033301.g001
Таким образом, нашей общей целью было исследовать сохранившуюся популяцию деревьев на предмет функциональных ветровых воздействий и количественно оценить масштабы этих эффектов на уровне лесного ландшафта. Поскольку влияние на рост выживших практически не учитывалось в предыдущих оценках ветровых воздействий в ландшафтном масштабе (например, [3]), мы проверили гипотезу о том, что краткосрочный рост выживших лесов не зависит от сильных ветров, с альтернативной гипотезой о том, что рост изменился. значительно за три года после урагана.С этой целью мы изучили лесной ландшафт площадью 6,79 млн га на юге Швеции, пострадавший от урагана Гудрун в январе 2005 г. Судя по структурному повреждению примерно 75 млн м 3 3 древесины (что составляет 110% от среднего ежегодный урожай 1998–2004 гг. в Швеции только с 16% лесной площади страны) Гудрун был самым сильным штормом в истории Швеции [19]. Тем не менее, этот серьезный ущерб пришелся на менее чем 6% [20] общей площади, пострадавшей от сильных ветров погодной системы Гудрун (для которой сообщалось о скорости порывов ветра до 42 м · с −1 , [3 ]).Здесь мы сосредотачиваемся на оставшихся 94% ландшафта, т. Е. На уцелевшей популяции деревьев, и исследуем изменения роста доминирующих видов деревьев (например, европейской ели, Picea abies (L.) Karst.) В 2005 г. –2007.
Результаты
Функции реагирования на изменение климата для контроля влияния изменения климата
Мы применили анализ функции отклика, чтобы исключить влияние изменения климата на наблюдаемые временные ряды роста, используя в качестве независимых переменных месячную температуру и количество осадков с июня предыдущего года по сентябрь текущего года.Климатические факторы объяснили 63,0% дисперсии роста деревьев в среднем по всем округам (Таблица 1). Этот уровень определенности находится в диапазоне, о котором сообщалось в предыдущих исследованиях по Скандинавии, согласно которым изменение климата определяет от 24% до 82% годового изменения роста деревьев [21] – [25]. Было установлено, что во всех округах температура и осадки в мае и июне текущего года оказали сильнейшее положительное влияние на прирост деревьев, но влияние отдельных климатических факторов значительно варьировалось между округами.Климатические факторы в предшествующий вегетационный период вместе составляли примерно 40% от общего влияния климата на рост деревьев [23], вывод, который впоследствии был использован для обоснования нашей гипотезы о временном характере функционального ветрового воздействия (см. «Материалы и методы»).
Исследовательский анализ роста до и после урагана
После учета влияния изменчивости климата на рост деревьев поисковый анализ выявил повсеместное снижение роста в годы после урагана (рис. 2).Прирост ели обыкновенной снизился во всех пострадавших от Гудрун округах по сравнению с уровнем до урагана. В течение трех лет после урагана уровни роста были ниже долгосрочного среднего в пяти из семи исследованных округов (в диапазоне от -8,0% до -0,7% ниже долгосрочного среднего). В этих же округах самые высокие скорости ветра были зарегистрированы во время Гудруна (рис. 2). Более того, в то время как аномалии роста ели обыкновенной (после учета влияния климата) в 2005–2007 гг. Показали положительную тенденцию для остальной части Швеции, они уменьшились в округах, затронутых Гудрун (Рисунок 3a, b).
Рис. 2. Аномалии роста деревьев до и после шторма Гудрун (январь 2005 г.) на юге Швеции.
Изображенные аномалии роста представляют собой отклонение от среднего долгосрочного ряда без возрастной тенденции после учета влияния климатических изменений. Столбцы показывают средние значения за три вегетационных периода до и после шторма. На вставленной карте показаны усредненные по площади максимальные скорости порывов ветра во время Гудруна [3]. Пространственные объекты – это группы округов, используемые Шведской национальной инвентаризацией лесов [27].U-S-S: Упсала, Стокхом и Седерманланд; Ö-V: Эребру и Вестманланд; S-Ä: Скараборг и Эльвсборг; Ö-K: Östergötland & Kalmar; JK: Jönköping & Kronoberg; GH: Гетеборг и Халланд; М-К-Б: Мальмохус, Кристианстад и Блекинге.
https://doi.org/10.1371/journal.pone.0033301.g002
Рисунок 3. Аномалии роста ели обыкновенной во времени.
На верхних панелях сравниваются аномалии роста (а) для округов, не затронутых Гудрун (например, Норрботтен-Лаппмарк, Норрботтен-Кустланд, Вестерботтен-Лаппмарк, Вестерботтен-Кустланд, Емтланд, Вестерноррланд, Евле, Вельмаланд и Даларланд) ) те, которые пострадали от урагана в январе 2005 г. (см. рис. 2).На нижних панелях приведены примеры временного развития наблюдаемых аномалий роста (кружки) для округов Йёнчёпинг и Кроноберг (c) и Мальмохус, Кристианстад и Блекинге (d) с подобранными моделями прерывистого временного ряда, обозначенными пунктирной линией. Сплошные горизонтальные линии на всех панелях обозначают среднюю аномалию роста за десять лет до урагана и в три года после Гудрун (серая заштрихованная область).
https://doi.org/10.1371/journal.pone.0033301.g003
Анализ временных рядов для проверки влияния ветра на рост
Для более систематического исследования функциональных ветровых воздействий и учета временных закономерностей и изменений в росте мы провели анализ прерывистых временных рядов, проверяя значимость и величину снижения ветрового роста в течение трех лет после урагана. Порядок процесса автокорреляции (то есть компонента AR) и процесса скользящего среднего (то есть компонента MA) в окончательно подобранных моделях находился в диапазоне от нуля до двух, соответственно.Окончательные модели ARIMA были несмещенными, а их остатки не имели значительной автокорреляции (таблица 1). Предполагаемая временная картина возмущения в целом хорошо согласуется с наблюдениями (см. Рис. 3c, d), а систематический анализ чувствительности фиктивных переменных возмущений показал, что средняя чувствительность оцененных коэффициентов возмущения по всем округам является умеренной (0,3 ± 2,0 процентных пункта).
Мы обнаружили, что снижение роста было значительным (α = 0,05) по сравнению с остаточной вариацией временного ряда роста в трех юго-западных округах исследуемого региона (Таблица 2).Кроме того, снижение роста во всех округах, кроме самых северных, оказалось незначительным, что примечательно, учитывая, что доступные временные ряды были относительно короткими (n = 56) по сравнению с рекомендациями для прерванного анализа ARIMA [26]. Таким образом, нам пришлось отвергнуть гипотезу о том, что Гудрун не повлияла на рост ели европейской. Оценки максимального снижения годового прироста, то есть коэффициенты возмущения анализа прерванных временных рядов, находились в диапазоне от -10.От 6% до 0,0% (Таблица 2), в среднем –6,2% по всей территории, затронутой Гудрун (с учетом объема стоячей перед ураганом).
Эти повсеместные функциональные эффекты были сильно связаны со скоростью ветра погодной системы Гудрун (рис. 4a). Линейная взаимосвязь между усредненными по площади максимальными скоростями порывов ветра и коэффициентами возмущения объясняет 85% вариации снижения роста и предполагает, что этот тип повреждения ветром происходит, если крупномасштабные максимальные скорости порывов превышают 25,5 м · с -1 .Взаимосвязь между структурными и функциональными эффектами шторма (рис. 4b) была немного менее сильной, но процент ущерба от ветровалов и разрушений все еще объяснял более трех четвертей наблюдаемых изменений в сокращении роста после урагана. Наши результаты показывают, что на каждые 10% объема древесины на корню, структурно поврежденной Гудрун, в среднем в течение трех лет после урагана наблюдалось сокращение прироста на 6,7%.
Рис. 4. Связь функционального ветрового воздействия с максимальной скоростью порыва ветра и повреждениями конструкции.
Панели показывают снижение роста, объясняемое максимальной скоростью порыва ветра (a) и гибелью деревьев от ветровоза и вырубки (b) для округов, пострадавших от урагана Гудрун. Функциональные ветровые эффекты представлены в виде ежегодных потерь прироста, усредненных за первые три вегетационных сезона после урагана, и были получены как коэффициенты возмущения в прерванном анализе ARIMA. Поскольку пересечение регрессии на панели (а) не было значимым, была подобрана модель без пересечения, в результате чего коэффициент регрессии был равен 0.672 (P <0,001). Максимальные скорости порывистого ветра на панели (а) представляют собой максимум за время шторма, пространственно усредненный по территории округа.
https://doi.org/10.1371/journal.pone.0033301.g004
Количественная оценка функционального ветрового воздействия в ландшафтном масштабе
Применяя оценочные коэффициенты возмущения к увеличенным значениям приращения Шведской национальной инвентаризации лесов [27], мы оцениваем, что ураган Гудрун вызвал общие потери прироста ели норвежской в 3 раза.0 млн. М 3 в период с 2005 по 2007 год. Только в округах Йёнчёпинг и Круноберг это сокращение превысило 1 млн. М 3 (Таблица 2). Таким образом, общая потеря роста в первые три года после урагана была больше, чем средний структурный ущерб от 77 ураганов, зарегистрированных в период с 1900 по 2004 год для Швеции, который, по подсчетам Нильссона и др., Составил 1,4 миллиона м. 3 . (2004). Кроме того, он был того же порядка величины, что и объем, поврежденный еловым короедом (т.е., основной вторичный агент возмущения, нанесенный ветром в экосистемах ели европейской) на юге Швеции в течение четырех лет после Гудрун [28]. Мы также можем сравнить потери прироста в результате единственного шторма Гудрун со среднегодовым ущербом континентального масштаба от короедов (т. Е. Наиболее вредоносного агента биотических нарушений в Европе), который оценивался в 2,9 млн. М 3 в период с 1950 г. и 2000 [1].
Обсуждение
Эффекты функциональных нарушений, такие как изменения роста после ветровых событий, до сих пор привлекали лишь ограниченное внимание в литературе (обратите внимание, например, на их отсутствие в основополагающем обзоре Эверхэма и Брокоу [12]).Кроме того, такие эффекты в настоящее время не учитываются при мониторинге и экономической оценке ущерба от ветра в контексте управления лесами, которые сосредоточены исключительно на структурных эффектах [1], [3], [29]. Хотя как положительные, так и отрицательные изменения роста, вызванные ветром, были зарегистрированы на уровне отдельных деревьев [11] – [18], мы показали здесь, что шторм Гудрун привел к повсеместному и значительному сокращению роста еловых лесов Норвегии на ландшафте. шкала. Значительные масштабы сокращения роста, задокументированные здесь, подчеркивают важность более целостного рассмотрения вызванных нарушениями изменений экофизиологических процессов при оценке воздействия нарушений на древесные ресурсы и лесное хозяйство (см. [30]).
Наши результаты, вероятно, также будут иметь значение для цикла C в лесу: Lindroth et al. [5] обнаружили глубокое изменение баланса углерода в лесу на участках, обрушенных штормом Гудрун. Поскольку рост ствола тесно связан с чистой продуктивностью экосистемы в спелых лесах [31], наши результаты показывают, что эффекты цикла углерода могут не ограничиваться участками, непосредственно поврежденными ураганом, но могут быть гораздо более широко распространенными в ландшафтах, затронутых сильными ветрами. Однако для точной количественной оценки этих эффектов необходимо исследование основных причин.Хотя наши данные подтверждают существование и значимость такого чистого ветрового эффекта в масштабе ландшафта, возникает вопрос, вызвано ли оно преимущественно снижением первичной продуктивности из-за затрудненного использования ресурсов (в результате механического повреждения корней, ксилемы и кроны) или из-за изменений в распределении (приоритезация подземных отсеков в ответ на механическое повреждение и воздействие ветра или выделение защитным соединениям для отражения атак насекомых) должно оставаться без ответа.
Результаты подробных эмпирических экспериментов в соседней Дании предполагают, что повреждение корней является вероятной причиной наблюдаемой потери роста: Нильсен и Кнудсен [14], проводя эксперименты по вытягиванию и высвобождению деревьев, обнаружили, что деревья европейской ели, подвергшиеся воздействию повышенной механической силы от ветра, пострадали от значительная потеря корня, связанная с повреждением корневой системы. Их образцы деревьев ответили последующей приоритизацией роста корней за счет верхних отделов ствола, что привело к пятилетнему увеличению базальной площади корней на 230%.В этом отношении анализ Vargas et al. [32] документально подтвердили, что деревья могут мобилизовать значительное количество накопленных углеводов, чтобы компенсировать потерю корней после урагана. Центральная роль повреждения корней в контексте снижения роста, о котором здесь сообщается, также подтверждается данными о повышенной чувствительности к засухе после Гудрун [33]. Эмпирические исследования, обнаружившие значительную потерю закрепления корней ели обыкновенной и увеличивающуюся склонность к последующему структурному повреждению ветром после Гудруна, также сообщают о прямой корреляции с показателями жизнеспособности деревьев и водного баланса [14], [34].
Поскольку как ветровые, так и засушливые возмущения будут независимо друг от друга предсказаны в будущем [35], [36], усиление обратных связей между ними может способствовать дальнейшему усилению режимов возмущений в условиях изменения климата (см. Также [37], [38]). Кроме того, поскольку рост и сила дерева связаны с защитой дерева от короедов [39], снижение продуктивности и повышенная подверженность засухе из-за повреждения корней могут быть основным механизмом, приводящим к широко наблюдаемым (например,г., [40]) повышение успешности колонизации и гибели короедов после шторма.
Что касается этих взаимодействий возмущений, важное ограничение в нашем понимании снижения роста после сильного ветра касается его продолжительности и устойчивости. Мы обнаружили значительные эффекты масштаба ландшафта для первых трех вегетационных периодов после Гудруна, но не можем сделать вывод о том, сколько времени займет крупномасштабное восстановление, исходя из наших данных. Басби и др. [41], например, сообщили о потере роста после урагана, который длился до пяти лет в отдельных насаждениях.Нильсен [34] обнаружил, что у двух третей исследованных деревьев по крайней мере 50% потери корней осталось через три года после шторма Гудрун. Таким образом, наши увеличенные результаты могут представлять собой первую консервативную оценку общей величины функциональных ветровых эффектов после Гудрун. Способность восстанавливаться до уровней роста, существовавших до нарушений, вероятно, также зависит от наличия дополнительных факторов стресса и общего состояния здоровья экосистемы. Кроме того, необходимо отметить, что в нашем анализе мы сосредоточили внимание исключительно на еловых лесах обыкновенной, и можно ожидать дифференцированных эффектов для видов с разной архитектурой корней или кроны, таких как сосна обыкновенная ( Pinus sylvestris L.), одного из основных представителей ели обыкновенной в изучаемом регионе.
В заключение, хотя возмущения существенно меняются в ответ на изменение климата и режимов управления [7], наше понимание их воздействия на лесные ресурсы и динамику экосистемы все еще остается неполным. В этом отношении значительный функциональный эффект ветра, о котором здесь сообщается, подчеркивает важность рассмотрения эффектов возмущения, помимо немедленной гибели деревьев, для разработки более целостного представления о том, как возмущения влияют на экосистемы [42], [43].Последнее крайне необходимо, так как природные нарушения все больше затрудняют устойчивое управление лесами, угрожая помешать достижению целей по увеличению использования возобновляемых ресурсов и смягчению последствий изменения климата посредством управления экосистемами [5], [44]. Принимая во внимание масштабы описанных здесь функциональных эффектов, мы делаем вывод, что влияние возмущения ветра на лесные ресурсы и углеродные балансы могло быть недооценено ранее, и призываем к более широкому рассмотрению воздействия возмущения на структуру экосистемы и , функционирующую в контексте функционирования управление лесным хозяйством и смягчение последствий изменения климата.
Материалы и методы
Данные
Переменной отклика в нашем анализе был рост ствола (т. Е. Косвенный показатель функционирования экосистемы, рис. 1) деревьев, переживших ураган Гудрун, полученный из кернов деревьев, взятых Шведской национальной инвентаризацией лесов (NFI). Все необходимые разрешения были получены для описанных полевых исследований (шведский закон 2001: 99). NFI систематически отбирает деревья по всей Швеции с целью получения репрезентативной информации о росте для всего диапазона насаждений и категорий владения на уровне округов (или их комбинаций, см. Рисунок 2).Годовые кольца анализируются NFI, т. Е. Перекрестным датированием, а возрастная тенденция удаляется стандартными процедурами анализа годичных колец [45]. Поскольку NFI ежегодно пересматривает только часть систематически выбранных мест отбора проб, мы использовали данные за четыре последовательных года инвентаризации (2007–2010) в нашем анализе. В целом, наш анализ основан на данных из 866 отдельных ядер деревьев (от 59 до 181 на исследуемый объект), которые были агрегированы для создания репрезентативных временных рядов роста для каждого округа за период с 1952 по 2007 годы.При анализе учитывались только ядра деревьев людей в возрасте старше 60 лет. Кроме того, мы сосредоточили внимание на ели европейской, которая является доминирующим видом в исследуемой области (доля от общего объема древесины на корню до урагана Гудрун: 46,6%, [27]), а также была наиболее пострадавшим видом от структурных повреждений урагана от Гудрун [20].
Чтобы устранить влияние климатических изменений на нашу переменную отклика (т. Е. Индекс годичных колец), мы провели анализ функции отклика (т. Е. Регрессию главных компонентов, см. [46], [47]), используя месячную температуру и осадки в качестве предикторов.Климатические данные за период с 1952 по 2007 год были получены из набора данных реанализа Kalnay et al. [48] (пространственное разрешение 2,5 ° × 2,5 °). Месячные переменные были центрированы и масштабированы по их среднему значению временного ряда и стандартному отклонению до их использования в анализе функции отклика. Основываясь на предыдущих выводах по Скандинавии [21], мы использовали 16 месяцев с июня предыдущего года по сентябрь текущего года в качестве потенциальных предикторов для индекса годичных колец. Ортогональность предикторов обеспечивалась проведением анализа главных компонент [49].Мы сохранили все основные компоненты в качестве предикторов в первоначальном регрессионном анализе [50], а затем выбрали лучшую и наиболее экономную модель путем обратного отбора, используя информационный критерий Акаике (AIC) в качестве показателя эффективности. Остатки выбранной модели (т. Е. Изменение индекса годичных колец, не связанное с изменением климата в анализе функции отклика) использовались в качестве переменной отклика в последующем анализе воздействия ветра на рост лесов.
Анализ прерванных временных рядов
После исследовательского анализа аномалий роста в вегетационные периоды до и после урагана мы провели анализ прерванных временных рядов, чтобы определить, оказало ли штормовое событие в январе 2005 г. значительное влияние на рост в следующие за ним вегетационные сезоны [51], [52] .Мы приспособили модели авторегрессионного интегрированного скользящего среднего (ARIMA) к 56-летним аномалиям роста, используя AIC для выбора модели. Параметр интегрирования ARIMA был установлен на ноль, поскольку никакого дрейфа не ожидалось из-за предварительного определения тенденции индекса древовидного кольца. Впоследствии мы включили в модель компонент возмущения, чтобы изучить влияние Гудруна на аномалии роста. Если предположить, что изменение во времени адекватно объясняется моделью ARIMA, значительное увеличение объяснительной силы за счет включения такого компонента возмущения будет указывать на то, что гипотеза об отсутствии влияния Гудрун на рост должна быть отклонена [51].Компонент возмущения, заданный как фиктивные переменные между 0 (т. Е. Отсутствие эффекта в годы до Гудруна) и 1 (т. Е. Максимальный эффект), был сформулирован как постепенный по началу и временный по продолжительности. Мы предположили, что функциональный эффект ветра достигнет максимума только во второй вегетационный период после урагана, учитывая временные задержки в экофизиологических корректировках деревьев (например, в отношении приоритетов распределения, см. [53]) и буферный эффект накопленных углеводов. в первый вегетационный период после нарушения.Gough et al. [54], например, обнаружили, что ежегодные наблюдения за фотосинтезом и ростом стебля были временно разделены из-за того, что фотосинтез в конце сезона был отнесен к росту стебля следующей весной (см. Также [23] и наши результаты по росту за предыдущий год в текущих условиях). годовой прирост, указанный выше). Кроме того, мы предположили, что восстановление эффекта начнется в третьем вегетационном периоде после ветрового события. Таким образом, фиктивные значения возмущений для первого и третьего вегетационного периода после шторма были установлены на 0.6 и 0,85 соответственно. Чтобы изучить чувствительность результатов к этой модели возмущений, мы дополнительно провели ряд анализов прерывистых временных рядов, варьируя фиктивные значения возмущений первого и третьего года от 0,5 до 1,0 с шагом 0,1 (всего 216 уникальных комбинаций). Наш анализ ограничивался первыми тремя вегетационными сезонами после урагана, поскольку достаточных данных о росте после 2007 года не было. Чтобы контролировать влияние второго шторма, обрушившегося на части исследуемой территории в 2007 году, мы ввели второй, отдельный коэффициент возмущения для этого года в соответствующих округах, чтобы отделить эффект этого второго возмущения от одного из Здесь анализируется шторм Гудрун 2005 года.Подходящие коэффициенты возмущения, т. Е. Влияние ветра на рост деревьев, определяемое прерванными моделями ARIMA, были проанализированы на предмет значимости и связаны с данными о скорости ветра, а также повреждении структурного шторма [3]. Важно отметить, что, поскольку в литературе сообщается как о положительных, так и отрицательных изменениях роста в ответ на сильный ветер на уровне отдельных деревьев [11] – [18], все анализы, оценивающие чистый эффект в масштабе округа, проводились как двусторонние тесты. .
Апскейлинг функциональных ветровых эффектов
Чтобы количественно оценить общий функциональный эффект Гудрун в масштабе ландшафта и связать его со структурным повреждением, мы масштабировали относительные изменения роста, определенные в анализе ARIMA, до абсолютных значений (т.е., м 3 ) для исследуемого ландшафта 6,79 · млн га. Этому масштабированию способствовало то, что мы основывали наш анализ на кернах деревьев NFI, то есть на записях деревьев из репрезентативной выборки еловых лесов на юге Швеции. Мы извлекли оценку роста ели обыкновенной в 2005–2007 годах из базы данных NFI ( i abs ) и оценили абсолютные изменения роста, вызванные ветром ( w abs ), исходя из относительных изменений роста. ( w отн. , i.е., коэффициенты возмущения ARIMA) согласно формуле. 1: (1) Все анализы проводились с использованием R Project for Statistical Computing v2.13.1 [55], в частности, с применением библиотечного прогноза [56] для моделирования ARIMA.
Благодарности
Мы благодарны Андерсу Лундстрему, SLU Umeå, за предоставление данных о росте NFI, и NOAA / OAR / ESRL PSD, Боулдер, Колорадо, США, за климатические данные повторного анализа NCEP. Кроме того, мы благодарим двух анонимных рецензентов за полезные комментарии к более ранней версии рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: KB. Проведены эксперименты: RS. Проанализированы данные: РС КБ. Предоставленные реагенты / материалы / инструменты анализа: KB. Написал статью: РС КБ.
Ссылки
- 1. Schelhaas MJ, Nabuurs GJ, Schuck A (2003) Природные нарушения в европейских лесах в XIX и XX веках. Биология глобальных изменений 9: 1620–1633.
- 2. Вестерлинг А.Л., Идальго Х.Г., Каян Д.Р., Светнам Т.В. (2006) Потепление и более раннее весеннее увеличение в западном U.С. лесная пожарная активность. Наука 313: 940–943.
- 3. Skogsstyrelsen (2006) Stormen 2005 – en skoglig analysis. Meddelande 1, 2006. Йёнчёпинг, Швеция. ISSN 1100-0295.
- 4. Курц В.А., Даймонд С.К., Стинсон Дж., Рэмпли Дж. Дж., Нейлсон и др. (2008) Кедровый жук и обратная связь углерода лесов с изменением климата. Nature 452: 987–990.
- 5. Линдрот А., Лагергрен Ф., Грелль А., Клемедссон Л., Лангвалл О. и др. (2009) Штормы могут вызвать сокращение поглотителя углерода в лесах в масштабах всей Европы.Биология глобальных изменений 15: 346–355.
- 6. Usbeck T, Wohlgemuth T., Dobbertin M, Pfister C, Bürgi A, et al. (2010) Увеличение ущерба лесам в Швейцарии от штормов с 1858 по 2007 гг. Сельскохозяйственная и лесная метеорология 150: 47–55.
- 7. Зейдл Р., Шелхас М.Дж., Лексер М.Дж. (2011a) Выявление движущих сил усиления режимов нарушения лесного покрова в Европе. Биология глобальных изменений 17: 2842–2852.
- 8. Зейдл Р., Шелхаас М.Дж., Линднер М., Лексер М.Дж. (2009) Моделирование нарушений, вызванных короедами, в крупномасштабной лесной сценарной модели для оценки воздействия изменения климата и оценки стратегий адаптивного управления.Региональные экологические изменения 9: 101–119.
- 9. Blennow K, Andersson M, Bergh J, Sallnäs O, Olofsson E (2010) Потенциальное изменение климата влияет на вероятность повреждения ветром лесов на юге Швеции. Изменение климата 99: 261–278.
- 10. Линднер М., Марошек М., Нетерер С., Кремер А., Барбати А. и др. (2010) Воздействие изменения климата, адаптивная способность и уязвимость европейских лесных экосистем. Экология и управление лесами 259: 698–709.
- 11.Мерренс Э.Дж., Пирт Д.Р. (1992) Влияние ущерба от урагана на индивидуальный рост и структуру древостоя в лиственных лесах в Нью-Гэмпшире, США. Журнал экологии 80: 787–795.
- 12. Everham EM, Brokaw NVL (1996) Ущерб лесам и восстановление после катастрофического ветра. Ботанический обзор 62: 113–185.
- 13. Zielonka T, Malcher P (2009) Динамика горного смешанного леса при ветровом возмущении в Татрах, Центральная Европа – дендроэкологическая реконструкция.Канадский журнал исследований леса 39: 2215–2223.
- 14. Nielsen CN, Knudsen MA (2004) Stormstabilitet og sundhed i en rødgranskærm. Данск Сковсбругс Тидсскрифт 89: 115–128.
- 15. Уэда М., Шибата Э. (2004) Почему деревья падают или умирают после сильного ветра? Состояние воды кипариса Хиноки, стоящего после тайфуна. Физиология деревьев 24: 701–706.
- 16. Coutand C, Dupraz C, Jaouen G, Ploquin S, Adam B (2009) Механические стимулы регулируют выделение биомассы в деревьях: демонстрация с молодыми деревьями Prunus avium.Анналы ботаники 101: 1421–1432.
- 17. Рубенс Б., Паннеманс Б., Данжон Ф., де Профт М., де Баэтс С. и др. (2009) Влияние механической стимуляции на развитие корней и побегов молодых контейнерных деревьев Quercus robur и Robinia pseudoacacia. Деревья 23: 1213–1228.
- 18. Hadley KS, Knapp PA (2011) Обнаружение сильных ветров с использованием данных годичных колец. Канадский журнал исследований леса 41: 1121–1129.
- 19. Nilsson C, Stjernquist I, Bärring L, Schlyter P, Jönsson AM и др.(2004) Зарегистрированные ураганы в шведских лесах 1901–2000 гг. Экология и управление лесами 199: 165–173.
- 20. Валингер Э., Фридман Дж. (2011) Факторы, влияющие на вероятность ветрового выброса на уровне древостоя в результате зимнего шторма Гудрун на юге Швеции. Экология и управление лесами 262: 398–403.
- 21. Linderholm HW (2002) Вариации роста сосны обыкновенной в XX веке в горах Центральной Скандинавии, связанные с изменением климата. Исследования Арктики, Антарктики и Альп 34: 440–449.
- 22. Linderholm HW, Moberg A, Grudd H (2002) Сосны на торфяниках как индикаторы климата? Региональное сравнение влияния климата на рост сосны обыкновенной в Швеции. Канадский журнал исследований леса 32: 1400–1410.
- 23. Selås V, Piovesan G, Adams JM, Bernabei M (2002) Климатические факторы, контролирующие воспроизводство и рост европейской ели на юге Норвегии. Канадский журнал исследований леса 32: 217–225.
- 24. Linderson H (2003) Сравнение ширины годичных колец современных древостоев сосны обыкновенной (Pinus sylvestris) и ели обыкновенной (Picea abies) с метеорологическими данными из двух районов Швеции – выводы для использования ширины годичных колец в качестве индикатора климата.Отчет Lundqua 38, Лундский университет, Лунд, Швеция. ISSN 0281-3076.
- 25. Grudd H (2008) Ширина и плотность годичных колец Торнетреска AD 500–2004: испытание на чувствительность к климату и новая 1500-летняя реконструкция лета в северной Фенноскандии. Климатическая динамика, 31: 843–857.
- 26. Box GEP, Jenkins GM (1970) Анализ временных рядов: прогнозирование и контроль. Сан-Франциско: Холден-Дэй.
- 27. NFI (2011) Статистика. Riksskogstaxering.Sveriges Lantbruksuniversitet. Доступно: http://www.slu.se/sv/webbtjanster-miljoanalys/statistik-om-skog/. По состоянию на 26 августа 2011 г.
- 28. Långström B, Lindelöw Å, Schroeder M, Björklund N, Öhrn P (2009) Вспышка елового короеда в Швеции после январских штормов 2005 и 2007 годов. Труды Рабочей группы IUFRO 7.03.10, Методология исследования лесных насекомых и болезней в Центральной Европе. 30 ноября 2009 г., Зволен, Словакия.
- 29. Nieuwenhuis M, O’Connor E (2001) Оценка финансового воздействия катастрофического ущерба от урагана в лесном хозяйстве Ирландии: тематическое исследование.II. Потеря доходов в будущем. Лесное хозяйство 74: 383–393.
- 30. Тернер М.Г. (2010) Нарушения и динамика ландшафта в меняющемся мире. Экология 91: 2833–2849.
- 31. Цвайфель Р., Эугстер В., Эцольд С., Доббертин М., Бухманн Н. и др. (2010) Связь между непрерывными изменениями радиуса ствола и чистой продуктивностью экосистемы субальпийского ельника норвежского в Швейцарских Альпах. Новый фитолог 187: 819–830.
- 32. Варгас Р., Трумбор С.Е., Аллен М.Ф. (2009) Доказательства использования старого углерода для выращивания новых тонких корней в тропическом лесу.Новый фитолог 182: 710–718.
- 33. Болте А., Хильбриг Л., Грундманн Б., Кампф Ф., Брюнет Дж. И др. (2010) Изменение климата влияет на структуру древостоя и конкурентные взаимодействия в елово-буковом лесу на юге Швеции. Европейский журнал исследований леса 129: 261–276.
- 34. Nielsen CN (2011) пытается найти простой динамизм и скрыть антагонизм. Сковен 3: 137–139.
- 35. Blennow K, Olofsson E (2008) Вероятность ветрового ущерба в лесном хозяйстве при изменении ветрового климата.Изменение климата 87: 347–360.
- 36. Уильямс А.П., Аллен С.Д., Миллар С.И., Светнам Т.В., Михаэльсен Дж. И др. (2010) Реакция лесов на повышение засушливости и тепла на юго-западе США. Труды Национальной академии наук Соединенных Штатов Америки 107: 21289–21294.
- 37. Фукуда К., Нишия Й., Накамура М., Судзуки К. (1997) Водные отношения ели Йезо и пихты Тодо в упавших насаждениях бореальных лесов на Хоккайдо, Япония. Журнал лесных исследований 2: 79–84.
- 38. Карникер Дж., Колл М., Ниньерола М., Понс Х, Санчес Дж. И др. (2011) Повсеместное ухудшение состояния кроны, разрушение пищевой сети и усиление смертности деревьев с усилением засухи, вызванной изменением климата. Труды Национальной академии наук Соединенных Штатов Америки 108: 1474–1478.
- 39. Кристиансен Э., Уоринг Р. Х., Берриман А. А. (1987) Устойчивость хвойных деревьев к нападению короеда: поиск общих взаимосвязей. Экология и управление лесами 22: 89–106.
- 40. Komonen A, Schroeder LM, Weslien J (2011) Развитие популяции Ips typographus после сильного шторма в заповеднике на юге Швеции. Журнал прикладной энтомологии 135: 132–141.
- 41. Busby PE, Motzkin G, Boose ER (2008) Изменения на уровне ландшафта в реакции леса на возмущение ураганом через тропу шторма. Канадский журнал исследований леса 38: 2942–2950.
- 42. Gutschick VP, BassiriRad H (2003) Экстремальные события как формирующие физиологию, экологию и эволюцию растений: к единому определению и оценке их последствий.Новый фитолог 160: 21–42.
- 43. Зайдл Р., Фернандес П.М., Фонсека Т.Ф., Жилле Ф., Йонссон А.М. и др. (2011b) Моделирование естественных нарушений в лесных экосистемах: обзор. Экологическое моделирование 222: 903–924.
- 44. Зейдл Р., Раммер В., Лексер М.Дж. (2011c) Уязвимость устойчивого лесопользования в Восточных Альпах к изменению климата. Изменение климата 106: 225–254.
- 45. Йонссон B (1969) Studier over den av väderleken orsakadeariations in årsringsbredderna hos tall och gran i Sverige.Rapporter och Uppsatser 16, Institutionen för Skogsproduktion, Стокгольм, Швеция.
- 46. Fritts HC (1976) Годовые кольца и климат. Academic Press, Нью-Йорк, Нью-Йорк, США. 567 с.
- 47. Fekedulegn BD, Colbert JJ, Hicks RR, Schuckers ME (2002) Как справиться с мультиколлинеарностью: пример применения регрессии главных компонентов в дендроэкологии. 43 с. Res. Пап. NE-721, Департамент сельского хозяйства, Лесная служба, Северо-Восточная научно-исследовательская станция.
- 48.Калнай Э., Канамицу М., Кистлер Р., Коллинз В., Девен Д. и др. (1996) 40-летний проект повторного анализа NCEP / NCAR. Бюллетень Американского метеорологического общества 77: 437–471.
- 49. Морцух Б.Дж., Руарк Г.А. (1991) Регрессия основных компонентов для смягчения эффектов мультиколлинеарности. Лесоводство 37: 191–199.
- 50. Jolliffe IT (1982) Примечание об использовании главных компонентов в регрессии. Прикладная статистика 31: 300–303.
- 51.McDowall D, McCleary R, Meidinger EE, Hay RA (1980) Анализ прерванных временных рядов. Беверли-Хиллз: Sage Publications.
- 52. Карпентер С.Р. (1990) Крупномасштабные возмущения: возможности для инноваций. Экология 71: 2038–2043.
- 53. Петрич Р., Хазенауэр Х., Пич С.А. (2007) Включение реакции роста лесов на прореживание в биоме-BGC. Экология и управление лесами 242: 324–336.
- 54. Gough CM, Vogel CS, Schmid HP, Su HB, Curtis PS (2008) Многолетняя конвергенция биометрических и метеорологических оценок накопления углерода в лесах.Сельскохозяйственная и лесная метеорология 148: 158–170.
- 55. R Development Core Team (2011) R: Язык и среда для статистических вычислений. R Фонд статистических вычислений, Вена, Австрия. ISBN 3-1-07-0. Доступно: http://www.R-project.org/. По состоянию на 26 августа 2011 г.
- 56. Hyndman RJ, Khandakar Y (2008) Автоматическое прогнозирование временных рядов: пакет прогнозов для R. Journal of Statistical Software 26: 3.
7-футовая искусственная елка из норвежской ели с предварительно зажженным или не освещенным шарниром – GDFStudio
Принесите своему дому атмосферу праздника в этот праздничный сезон с деревом, которое придаст вашему интерьеру очарование очаровательного леса.С нашим неосвещенным или предварительно освещенным искусственным деревом вы сможете создать воспоминания на всю жизнь, не беспокоясь о том, чтобы убирать иголки или поддерживать свое дерево в живых каждый год. Этот аксессуар идеально моделирует элегантную норвежскую ель, придавая вашим украшениям пышный зеленый вид, а также встроенные фонари, которые освещают каждый праздничный вечер уютным теплым светом.
НОРВЕГИЯ ЕЛКА: Наше дерево от темно-зеленого цвета до элегантной конической формы идеально передает очарование изысканной елки из норвежской ели.Это дерево, известное своими закругленными иглами, обращенными вперед, имеет пышные кончики и мутовчатые ветви, которые идеально подходят для подвешивания украшений. Обратите внимание, что этот аксессуар предназначен только для использования в помещении.
НЕ ОСВЕЩЕННОЕ ИЛИ ПРЕДВАРИТЕЛЬНОЕ ОСВЕЩЕНИЕ: вы можете выбрать между неосвещенным деревом и предварительно освещенным деревом, в котором используются обычные струнные огни. Наше неосвещенное дерево позволяет вам строить пустой холст, в то время как наш вариант струнного освещения бывает как прозрачным, так и многоцветным, чтобы предложить более традиционный вид.
ПОДСТАВКА В КОМПЛЕКТЕ: Наше дерево поставляется с прочной железной подставкой, гарантирующей, что ваше дерево будет оставаться устойчивым и закрепленным во время использования.Это закончено с накладками для ног, чтобы предотвратить любые царапины или порезы на полу.
РАЗМЕРЫ: Выберите украшение для праздника, которое идеально подходит по размеру для вас и вашего дома. Эта рождественская елка имеет размеры 58,00 “Ш x 58,00” x 84,00 “В с основанием 27,50” Ш x 27,50 “Г x 7,75” В. Вам понравится, насколько праздничным может стать ваше внутреннее пространство с простым добавлением этого очаровательного дерева.
ТРЕБУЕТСЯ СБОРКА: Для этой елки требуется некоторая сборка.В комплект входят все инструкции и инструменты, необходимые для сборки.
- Включает: 1 (одну) искусственную новогоднюю елку
- Материал иглы: ПВХ
- Породы деревьев: Ель обыкновенная
- Конструкция: навесная
- Цвет иглы: зеленый
- Цвет света: не горит / многоцветный / прозрачный
- Количество ламп: 700
- Количество ответвлений: 2231
- Подставка в комплекте
- Огнестойкий
- Детали ручной работы
- Требуется некоторая сборка
- Размеры: 58.00 дюймов в глубину x 58,00 дюймов в ширину x 84,00 дюймов в высоту
- Размеры основания: 27,50 дюйма в глубину, 27,50 дюйма в ширину и 7,75 дюйма в высоту
Маленькая жемчужина норвежской ели для продажи онлайн
Ель Little Gem – чудесное растение и замечательный подарок. Он образует идеальную шаровидную вечнозеленую форму насыщенного зеленого цвета на прямостоячем стволе. Чудо в том, что он от природы карликовый и никогда не нуждается в обрезке, но всегда выглядит идеально сферическим и аккуратным. Это идеальный ответ для тех, кто хочет добавить формальности своему саду, например, у входа или вдоль дорожки, но боится всех обычно требуемых стрижек.Это дерево, которое будет расти в холодных регионах до минус 30 градусов, а также хорошо расти в теплых и жарких регионах. Для наилучшего роста его следует высаживать на открытом солнце, а в очень жаркую погоду следует укрыться в полдень. Он легко растет на большинстве почв и устойчив к засухе при укоренении.
- Идеальный способ придать официальный вид без необходимости закреплять и закреплять
- Яркие желто-зеленые молодые побеги весной
- Растет на любом солнечном месте и на любой почве
- Великолепное вечнозеленое растение насыщенно-зеленого цвета, создающее потрясающий акцент.
- Безотказный, морозоустойчивый и засухоустойчивый
Для создания великолепного формального образа без каких-либо дополнительных усилий, ель Little Gem должна быть выбором номер один.Это дерево станет отличным подарком для тех, кто любит формальность, но ведет жизнь, слишком занятую для всей этой стрижки. Подумайте о том, как идеально они будут выглядеть парой по обе стороны от дверного проема, чтобы придать особый вид любому дому.
Сады с более формальным оформлением определенно вернулись в моду, и формальные штрихи также добавляют много к более непринужденному стилю садоводства. Одна из широко используемых формальных деталей – это обрезанный круглый шар на вертикальном стволе, который можно использовать для акцентирования углов, создания линий или для укладки в ящики рядом с дверями и ступенями.Единственная проблема, связанная с этой функцией, заключается в том, что шары нужно тщательно обрезать и заботиться о том, чтобы они выглядели аккуратно и элегантно, что очень важно для правильного «внешнего вида». Таким образом, идеальное растение для этого формального использования никогда не потребует стрижки. Но эй, такого не может быть, правда? Без ошибок! Вы можете отправить кому-нибудь в подарок – может быть, себе – идеальное круглое растение, которое никогда не будет нуждаться в стрижке и всегда будет выглядеть аккуратно. Это ель Little Gem.
Сейчас ель для большинства людей звучит как большое лесное дерево, и это правильно.А вот Little Gem Spruce – полная противоположность. Это одно из чудес природы; плотное крошечное растение, образующее идеальную сферу, не более 2 футов в поперечнике . Производство таких особенных деревьев требует квалифицированной работы, поэтому, к сожалению, их всегда не хватает, а зачастую и вовсе нет.
Growing Little Gem Норвежские ели
Итак, представьте – растение, которое без стрижки естественным образом вырастет в шар, который никогда не станет слишком большим. Наши опытные садоводы установили это удивительное дерево на прочный ствол, так что это идеальное «леденцовое дерево» – сферический шар на стволе, который никогда не нуждается в обрезке, но всегда выглядит аккуратно. .Это дерево образует густую группу из коротких стеблей, покрытых короткими иголками, причем стебли так плотно прилегают друг к другу, что образуют сплошной шар. Скорость роста очень равномерная, поэтому мяч просто медленно становится больше, растет во всех направлениях с одинаковой скоростью и никогда не дает длинных побегов, которые нужно обрезать. Весной новообразование желто-зеленого цвета, почти как цветущий шар. С наступлением лета стебли становятся темно-зелеными и остаются такими же всю зиму.
Выносливость
Маленькая самоцветная ель очень вынослива, и будет расти в каждом штате по всей стране.Это особенно полезно в более холодных регионах, где выбор растений более ограничен. Его следует сажать на открытом солнце в более прохладных местах и в полутени в жарких регионах, где полезно укрыться от полуденного солнца. Это дерево хорошо работает на большинстве типов почв, кроме резко щелочных, и оно будет расти легко, без суеты и беспокойства. После посадки его следует хорошо поливать, особенно в первое лето, но после укоренения он без проблем переживет периоды засухи.
Используется в вашем саду
Используйте ель Little Gem, где бы вы ни хотели устроить торжественный бал. Это может быть по обе стороны от входа, прямо в земле или в паре красивых горшков. Они могут обозначать углы клумбы или указывать высоту над клумбой или горшком с цветущими растениями или низкорослыми кустарниками под ними. Используйте несколько, чтобы обозначить дорожку или придать структуру любой части сада. Поместите их в круг вокруг фонтана. Это идеальное растение везде, где требуется формальный подход, потому что никому не нужно постоянно находиться в саду, подстригая непослушное растение, – природа сделает это за вас, и это дерево никогда не будет выглядеть неухоженным.
История и происхождение маленького драгоценного камня обыкновенной ели
У этого дерева интересная история. Ель обыкновенная ( Picea abies ) естественным образом растет в Скандинавии, на западе России и в горах Европы. Несмотря на то, что это красивое дерево, оно не обладает достаточными особенностями, чтобы выращивать его в садах, но его выращивают на больших плантациях в качестве источника пиломатериалов из хвойных пород. Очень редко с некоторыми деревьями случаются странные вещи. Из ветки вырастет крошечная гроздь карликовых ветвей, которая называется «ведьмин метла».Из кусочков этих крошечных ветвей, если их правильно выращивать, вырастут крошечные растения, и несколько карликовых вечнозеленых растений зародились именно так.
Примерно в начале -х годов века в питомнике недалеко от Гамбурга, Германия, это произошло на ели европейской, и полученное в результате растение, названное ель Птичьего гнезда ( P. abies ‘Nidiformis’), представляет собой очень популярное и широко выращиваемое карликовое вечнозеленое растение до сих пор. Однако, несмотря на то, что он карликовый, он не такой уж и маленький, и многие садовники были удивлены, когда этот милый маленький кустик быстро превратился в растение 4 фута в высоту и 6 футов в поперечнике – или даже больше.