архитектурный элемент – это… Что такое архитектурный элемент?
- архитектурный элемент
арка – криволинейное перекрытие пролета.
аркада.
интерколумний. | аркбутан.
курватура.
контрфорс.
Идеографический словарь русского языка. — М.: Издательство ЭТС. Баранов О.С.. 1995.
- колонна
- скульптурное украшение
Смотреть что такое “архитектурный элемент” в других словарях:
архитектурный элемент плоскости — фасад передняя сторона здания. тимпан. анты. ниша. альков. экседра. фонарь. эркер остекленная часть здания, выступающая за пределы плоскости фасада. аттик. фриз. фронтон вимперг. щипец … Идеографический словарь русского языка
Шатёр (архитектурный элемент) — Шатёр в деревянной архитектуре.

Астрагал архитектурный элемент — см. Архитектура и Приложение, таблица I, черт. 1 й … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Астрагал, архитектурный элемент — см. Архитектура и Приложение, таблица I, черт. 1 й … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Сима (архитектурный термин) — У этого термина существуют и другие значения, см. Сима (значения). Сима (др. греч. Σιμός курносый, выпуклый) элемент карниза, желобок для стока воды. Симы впервые появились в древнегреческой архитектуре во время господства дорийского ордера. Они… … Википедия
колонна
— Архитектурный элемент, играющий роль опорного столба или подражающий ему. Ист.: Плужников, 1995 • • • (лат. columna круглый столб). Архитектурно обработанный столб, несущий вертикальную нагрузку. В классических ордерах состоит из капители, ствола … Словарь храмового зодчестваЕвро — (Euro) Евро это единая европейская валюта Евро: описание монет и банкнот, история создания и развития, место в мировой экономике Содержание >>>>>>>>>> … Энциклопедия инвестора
Царицыно (дворцово-парковый ансамбль) — У этого термина существуют и другие значения, см. Царицыно. памятник архитектуры (федеральный) Дворцово парковый ансамбль … Википедия
Здание Главного штаба (Санкт-Петербург) — Здание Главного штаба … Википедия
Памятники А. С. Пушкину в Твери — А. С. Пушкин проезжал через Тверь почти тридцать раз, из них четырежды останавливался в городе[1]. Здесь с поэтом произошли некоторые знаменательные события[2], в том числе несостоявшаяся дуэль с В.
А. Соллогубом[3]. Тверь… … Википедия

 Ист.: Плужников, 1995 • • • (лат. columna круглый столб). Архитектурно обработанный столб, несущий вертикальную нагрузку. В классических ордерах состоит из капители, ствола … Словарь храмового зодчества
Ист.: Плужников, 1995 • • • (лат. columna круглый столб). Архитектурно обработанный столб, несущий вертикальную нагрузку. В классических ордерах состоит из капители, ствола … Словарь храмового зодчества А. Соллогубом[3]. Тверь… … Википедия
А. Соллогубом[3]. Тверь… … ВикипедияДекоративные элементы на фасаде – наименования
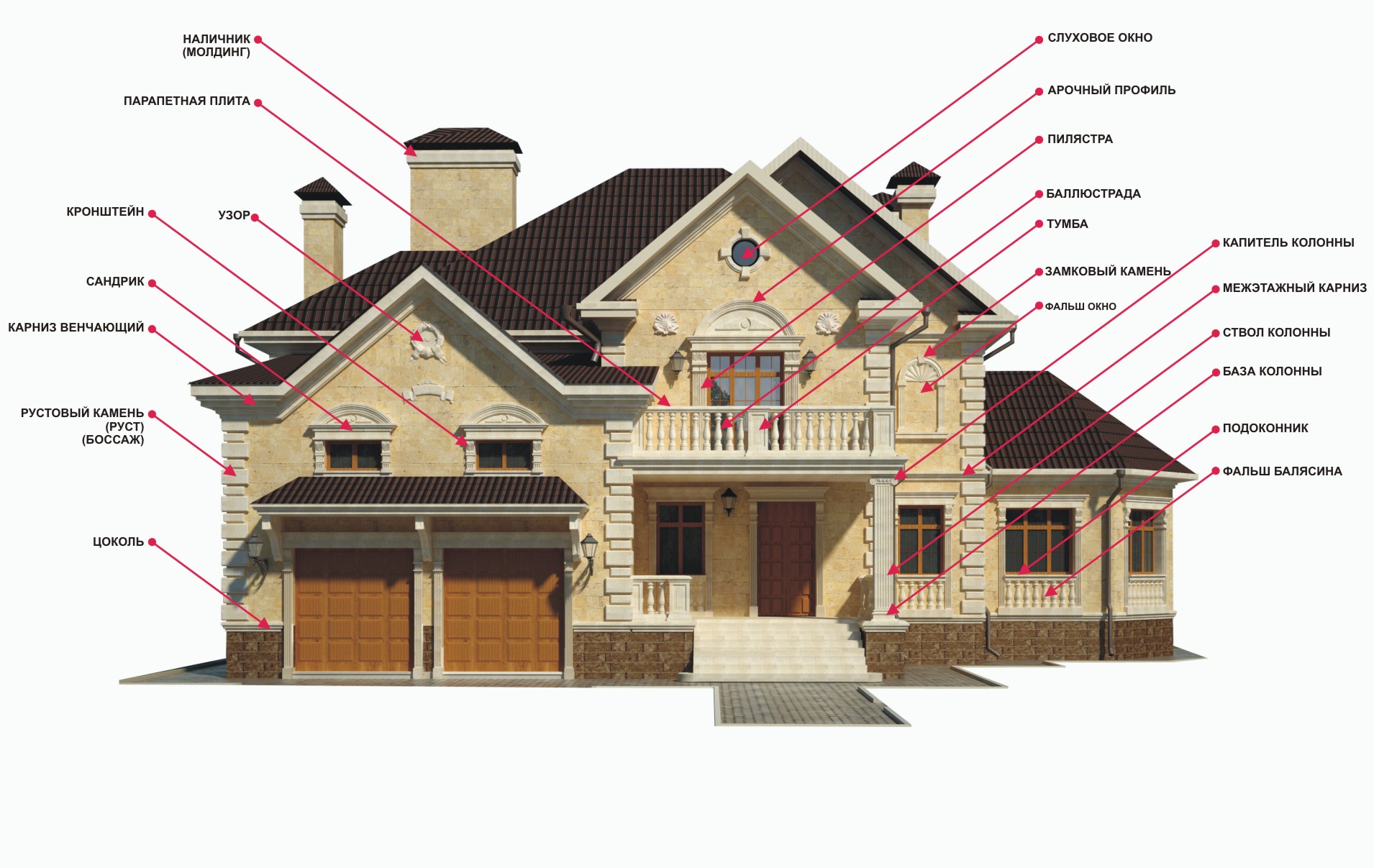
Декоративные обрамления на фасаде, или как правильно называется «вот такая штучка».
Давайте разберемся в фасадном декоре.
Какой он бывает? Как его назвать? И как правильно выразить свои мысли.
Аттик
Декоративная стенка, построенная над венчающим архитектурное строение карнизом. Чаще всего украшена рельефными надписями. Прямоугольное завершение Портика.
Арка
 0001pt;”>
0001pt;”>
Криволинейное перекрытие в пролете между двумя опорами (устоями моста или колонами). Создающее боковой распор.
По форме арки бывают:
– «полуциркульные» – с сечением полуокружности;
– «лучковые» – с дугой менее полуокружности;
– «стрельчатые» – образованные двумя арками, которые пересекаются под углом;
– «многолопастные» – сочетают в себе много мелких арочек.
Верхняя, несомая часть архитектурного ордена – балочное перекрытие пролета или завершения стен. Горизонтальная часть строения, которая опирается на колоны и состоит из карниза или фриза.
Антаблемент совместно с системой колон очень редко встречается за пределами классической архитектуры. Используется при отсутствии колон для завершения верхней части стены. Бывает, что антаблемент монтируют без его некоторых частей, например антаблемент без архитрава называется облегченным, а без фриза – неполным.
Используется при отсутствии колон для завершения верхней части стены. Бывает, что антаблемент монтируют без его некоторых частей, например антаблемент без архитрава называется облегченным, а без фриза – неполным.
Балюстрада
Балюстрада – это не высокое ограждение террасы, лестницы или балкона. Состоят из фигурных столбиков, соединенные перилами или горизонтальной балкой с верху. Предназначена для ограждения конструкций сложных форм, таких как круглые балконы, винтовые лестницы и т.д.
Болевер
Своеобразная надстройка над зданием, смотровая вышка, беседка. Чаще всего круглая в плане. Поставлена – для вида из него, так сказать для обзора местности.
Чаще всего круглая в плане. Поставлена – для вида из него, так сказать для обзора местности.
Балясины
Навесные фигурные столбики, иногда с резным декором. Поддерживающие перила ограждения балкона или лестницы и пр.
Волюта
Архитектурный элемент – спиралевидного завитка, с «глазком» в центре. Состоящая из ордерных капителей, архитектурная часть дверей, окон, карнизов и т.п.
Гирлянда
 85pt 0.0001pt;”>
85pt 0.0001pt;”>
Орнаментальный мотив из волнообразно свисающих лент, цветов, листьев и т.п. Такой вид декоративных обрамлений характерный для стиля барокко.
Галерея (гульбище)
Крытое, длинное строение, примыкающее к стене здания. Одну из стен которой заменяют колонны или столбы примыкающие к стене строения.
Гребень
Фигурные завитки, Служат для украшения верха стены.
Килеобразное или полукруглое завершение части наружных стен. Повторяет расположенного за ней свода очертания.
Кариатида
Опора в виде скульптурного изображения стоящей женской фигуры. Поддерживающая балочное перекрытии.
Капитель
Верхняя часть колонны или пилястра, в архитектуре служит переходом к покрытию, воспринимает нагрузку от горизонтальных балок перекрытия.
Карниз
В наружной отделке фасадов – выступающий элемент. Который разделяет горизонтальную или вертикальную плоскость по характерным линиям которое необходимо выделить.
Каннелюры
Каннелюры в архитектуре – вертикальные желобки на стволе пилястры или колонны.
Колона
Часть архитектурного сооружения в виде высокого столба, который в свою очередь служит несущей опорой фронтонов или внутренней части строения. Состоит из ствола, базы и капителей.
Состоит из ствола, базы и капителей.
Ствол покоится на простой или сложной базе, увенчанный капителью. Колоны применяются как для фасадной части зданий, так и для внутренней отделки. Отдельно стоящие колоны, украшенные скульптурой, чаше всего служат памятником. В каркасных домах, колоны чаще всего служат опорой, воспринимая на себя нагрузку от закреплённых к ним элементов.
База – основание, нижняя часть пилястры или колоны.
Консоль
В архитектурном строительстве – выступающий из стены элемент. Служащая, как для декора, так и в качестве полочки, на которую устанавливаются различные украшения, вазы и пр.
 85pt 0.0001pt;”>
Лепнина
85pt 0.0001pt;”>
Лепнина
Лепное, фигурное, рельефное украшение на фасадах и в интерьерах. Чаще всего отлитые или спресовынные из бетона, гипса и т.д.
Модильон
Модильон, еще используется название Модульон – украшение в карнизе, в виде небольшого кронштейна, с лепным листом в нижней части.
Наличник
Декоративное обрамление дверных и оконных проемов. Состоящий из фронтона. Так сказать накладная планка в виде рамы вокруг окон и дверей.
Ниша
Своеобразный архитектурный элемент, а попросту – углубление в стене дома, для установки различных предметов интерьера.
Экседрами называются полукруглые ниши.
Панно
Скульптурное или живописное изображение декоративного или тематического характера. Прикрепляются к стенам, потолкам и т.д., и обрамлено рамкой или лентой арнамента. Панно служит для постоянного или временного украшения
Парапет
 85pt 0.0001pt;”>
85pt 0.0001pt;”>
Не высокая сплошная стенка, которая проходит по краю террасы, балкона, крыши и т.д., на набережной в качестве заграждения, дамбы, в качестве защиты от действия волн. Также парапет служит так называемым постаментом, например для декоративных статуй.
Пилон
Колонна или столб, чаще всего со скульптурными украшениями, у входа в здание, в воротах зданий и т.д. служащие опорой перекрытий или отдельно стоящие по сторонам входов или въездов. Большие столбы служат опорой например в подземных станциях метро, либо поддерживают тросы на больших висящих мостах.
Портик
 85pt 0.0001pt;”>
85pt 0.0001pt;”>
Открытая галерея на колонах или столбах и помещенных перед фасадом здания. Обычно перед входом в здание, завершенная фронтоном или артиком.
Пилястра
Выступ, плоский или вертикальный, прямоугольного сечения на поверхности столба или стены. Пилястра обладает теми же частями (ствол, капитель, база), что и колона, а служит для деления плоскости стены. Служит преимущественно в качестве декоративного элемента. Иногда пилястра усиливает стену.
Рокайль
Главный элемент орнамента стиля рококо. Напоминает форму завитка раковины. В начале XXIII века появился во Франции, использовались при украшении павильонов-гротов деталями, имитирующими природные элементы (причудливые растения, морские раковины, обломки скал и т.д.). Иногда термин «Рокайль» применяется для обозначения стиля рококо.
В начале XXIII века появился во Франции, использовались при украшении павильонов-гротов деталями, имитирующими природные элементы (причудливые растения, морские раковины, обломки скал и т.д.). Иногда термин «Рокайль» применяется для обозначения стиля рококо.
Ротонда
Круглая в плане постройка, центрическое сооружение, чаще всего увенчанная куполом (храм, мавзолей, повильен).
Руст, рустовка, рустик
Рельефная кладка или облицовка стен камнем с выпуклой и грубо отесанной поверхностью. Создавая впечатление массивности и мощи строения. При отделке фасада штукатурками, рустик имитируется разбивкой стены на полосы и прямоугольники.
При отделке фасада штукатурками, рустик имитируется разбивкой стены на полосы и прямоугольники.
Сандрик
Декоративный архитектурный элемент в виде небольшого карниза, который располагается над проемом двери или окон на фасаде дома, иногда применяется в интерьере.
Сандрик иногда опирается на консоли и завершается фронтоном.
Тимпан
Углубленная часть стены, ниша, треугольного или стрельчатого очертания над дверьми или окнами. В Тимпан чаше всего размещают живописные картины, гербы, скульптуры и др.
Фасад
 85pt 0.0001pt;”>
Наружная сторона сооружения, здания, дома. В зависимости от формы строения и ее окружения различают
85pt 0.0001pt;”>
Наружная сторона сооружения, здания, дома. В зависимости от формы строения и ее окружения различают
-
главный фасад,
-
уличный фасад,
-
боковые фасады,
-
уличный,
-
дворовый,
-
парковый и другие фасады.
Пропорции, тектоническое и декоративное деления фасада как правило определены назначением здания, особенностями его стилистического, пространственного и конструктивного решения.
Фриз
Средняя горизонтальная часть антаблемента, между архитравом и карнизом. в дорическом ордере расчленяется на триглифы и метопы (триглифно-метопный фриз), в ионическом и коринфском ордерах заполняется сплошной лентой рельефов или оставляется пустым; сплошная полоса декоративных, скульптурных, живописных и других окаймляющая верх стен, поверхность пола помещения, поле ковра и др.
в дорическом ордере расчленяется на триглифы и метопы (триглифно-метопный фриз), в ионическом и коринфском ордерах заполняется сплошной лентой рельефов или оставляется пустым; сплошная полоса декоративных, скульптурных, живописных и других окаймляющая верх стен, поверхность пола помещения, поле ковра и др.
Фронтон
Лицевая часть стены, завершение фасада дома, портика, колоннады, ограниченное двумя скатами по бокам и карнизом у основания фронтона (тимпан) часто украшается скульптурой. Декоративные фронтоны украшают двери и окна зданий.
Цоколь
Цоколь – нижняя часть наружной стены здания, обычно несколько выступающая, сооружения, памятника или колонны, лежащая на фундаменте, обычно несколько выступающая. Цоколь обрабатывается рустовкой, профилями получает декоративную обработку.
Цоколь обрабатывается рустовкой, профилями получает декоративную обработку.
Энтазис
Энтазис – утолщение ствола(фуста) колонны в средней его части как правило на одну треть ее высоты, создающее впечатление напряженности и устраняющее оптическую иллюзию вогнутости ствола.
В данной статье мы очень коротко попытались объяснить основные архитектурные «словечки»
Вот теперь мы знаем все!
Читайте также:
Стоимости работ по монтажу декоративных элементов Вы можете найти в нашем разделе «цены»
Карамон — архитектурный элемент храмов
Карамон — ворота в китайском стиле, расположенные на территории храма Ниси Хонган-дзи. Были перенесены из замка Фусими. Ворота Карамон появились в эпоху Хэйан (8-12 век). В название ворот используется иероглиф кара, который означает «Китай» или «Тан» (название одной из династии в Китае). Тем не менее, изначально это слово означало «благородство» и «элегантность». В качестве приставки иероглиф также добавлялся к названиям объектов, которые считались величественными или замысловатыми, невзирая на их происхождение. В результате, ворота стали называться Карамон, поскольку были воплощением этих качеств.
Были перенесены из замка Фусими. Ворота Карамон появились в эпоху Хэйан (8-12 век). В название ворот используется иероглиф кара, который означает «Китай» или «Тан» (название одной из династии в Китае). Тем не менее, изначально это слово означало «благородство» и «элегантность». В качестве приставки иероглиф также добавлялся к названиям объектов, которые считались величественными или замысловатыми, невзирая на их происхождение. В результате, ворота стали называться Карамон, поскольку были воплощением этих качеств.
| Ворота Карамон в храме Ниси Хонган-дзи | фото: nimame |
Архитектурный стиль Карамон
Изображения Карамон можно часто увидеть на свитках эпохи Хэйан. Таким образом, Карамон — это не особое название ворот в храме Ниси Хонган-дзи, а разновидности ворот, которые обладают некоторыми отличительными архитектурными элементами. Первой особенностью Карамон было наличие карахафу — волнообразные доски на фронтонах ворот, украшенные резьбой, что является редким декоративным элементом для японской архитектуры. Во-вторых, у ворот четыре «ноги», в связи с этим они иногда также называются сикякумон. Дословно переводится как «ворота с четырьмя ногами». Но, даже не вдаваясь в архитектурные тонкости, видно, что Карамон очень сильно отличается от традиционного японского искусства, которому свойственна сдержанность и стремление к естественности. Декор же ворот поражает своим богатством, обилием всевозможных украшений и золота, яркостью красок.
Первой особенностью Карамон было наличие карахафу — волнообразные доски на фронтонах ворот, украшенные резьбой, что является редким декоративным элементом для японской архитектуры. Во-вторых, у ворот четыре «ноги», в связи с этим они иногда также называются сикякумон. Дословно переводится как «ворота с четырьмя ногами». Но, даже не вдаваясь в архитектурные тонкости, видно, что Карамон очень сильно отличается от традиционного японского искусства, которому свойственна сдержанность и стремление к естественности. Декор же ворот поражает своим богатством, обилием всевозможных украшений и золота, яркостью красок.
| Ворота Карамон в храме Ниси Хонган-дзи | фото: Ronald Woan |
Историческое значение Карамон
Изначально декоративный элемент карахафу использовался только в храмах и при входе в дома аристократии, но с приходом периода Адзути-Момояма (1568-1603) ворота Карамон стали важным архитектурным и декоративным элементом при строительстве домов даймё и замков. Карамон стояли на входе в поместье и использовались только при посещение сёгуном своих вассалов, а также при посещение императором поместья сёгуна. Благодаря этой традиции ворота Карамон приобрели особое значение и стали своеобразным воплощением социальной связи между различными слоями общества. В более поздний период ворота Карамон несколько изменили свое значение. Все больше стали символизировать престиж и важность строения, частью которого они были. Причем это было актуально как для религиозных, так и светских строений. С приходом эпохи Токугава (1603-1868) за воротами Карамон окончательно закрепилось значение власти и авторитета, отраженных в архитектуре.
Карамон стояли на входе в поместье и использовались только при посещение сёгуном своих вассалов, а также при посещение императором поместья сёгуна. Благодаря этой традиции ворота Карамон приобрели особое значение и стали своеобразным воплощением социальной связи между различными слоями общества. В более поздний период ворота Карамон несколько изменили свое значение. Все больше стали символизировать престиж и важность строения, частью которого они были. Причем это было актуально как для религиозных, так и светских строений. С приходом эпохи Токугава (1603-1868) за воротами Карамон окончательно закрепилось значение власти и авторитета, отраженных в архитектуре.
Ворота Карамон в Ниси Хонган-дзи были созданы в неспокойный период войн. Они воплощают собой силу и могущество великого полководца и объединителя Японии Тоётоми Хидэёси. На фронтонах ворот можно видеть герб рода Тоётоми — декоративный цветок павлонии, украшения, выполненные лучшими мастерами резьбы по дереву.
Эти ворота — один из немногих уцелевших элементов первоначального строения некогда прекрасного замка Фусими, сыгравшего столь важную роль в эпохе Сэнгоку. С 1994 года ворота Карамон в Ниси Хонган-дзи включены в список наследия ЮНEСКО вместе с основным комплексом храма.
С 1994 года ворота Карамон в Ниси Хонган-дзи включены в список наследия ЮНEСКО вместе с основным комплексом храма.
Советы гида по Киото самостоятельным туристам
Посещая храм Ниси Хонган-дзи, обязательно обойдите основной комплекс слева и посмотрите на ворота Карамон. Это единственное из трех сокровищ храма, для посещения которого не нужно специально бронировать экскурсию.
Видео о храме Ниси Хонган-дзи
виза в Японию | закажите дополнительно экскурсии в Японии | туры в Японию
Не удается найти страницу | Autodesk Knowledge Network
(* {{l10n_strings.REQUIRED_FIELD}})
{{l10n_strings.CREATE_NEW_COLLECTION}}*
{{l10n_strings.ADD_COLLECTION_DESCRIPTION}}
{{l10n_strings. COLLECTION_DESCRIPTION}}
{{addToCollection.description.length}}/500
{{l10n_strings.TAGS}}
{{$item}}
{{l10n_strings.PRODUCTS}}
{{l10n_strings.DRAG_TEXT}}
COLLECTION_DESCRIPTION}}
{{addToCollection.description.length}}/500
{{l10n_strings.TAGS}}
{{$item}}
{{l10n_strings.PRODUCTS}}
{{l10n_strings.DRAG_TEXT}}
{{l10n_strings.DRAG_TEXT_HELP}}
{{l10n_strings.LANGUAGE}} {{$select.selected.display}}{{article.content_lang.display}}
{{l10n_strings. AUTHOR}}
AUTHOR}}
{{l10n_strings.AUTHOR_TOOLTIP_TEXT}}
{{$select.selected.display}} {{l10n_strings.CREATE_AND_ADD_TO_COLLECTION_MODAL_BUTTON}} {{l10n_strings.CREATE_A_COLLECTION_ERROR}}Лестниц технологического парка Actiu, уникальный архитектурный элемент
Всемирный день архитектуры прошел 7 октября года, и за это время многие города по всему миру организовали недели, наполненные мероприятиями, связанными с архитектурой. В ознаменование этого важного события и разделяя приверженность Actiu архитектуре, мы хотели бы пригласить вас познакомиться с двумя архитектурными элементами , отличающими его Технологический парк: двумя лестницами.
Первый из них находится у входа в корпоративное здание по адресу Actiu Technology Park . Это здание вдохновлено основным принципом: создать наилучшие условия для работы. Для этого здание представляет собой открытое, светлое и комфортное здание, а также дизайн, эффективность и функциональность – три качества, которые Actiu демонстрирует во всех своих продуктах.
Лестница в холле корпоративного здания – первая крупная выставка дизайна и функциональности, которую мы находим.Этот шлюз дает нам доступ на первый этаж офисов компании. В этом архитектор парка, Хосе Мария Томас Ллавадор , хотел проиллюстрировать архитектурное предложение технологического парка Actiu в целом: риск, инновации и авангард. Эта лестница из ломаных линий, почти невозможная и очень чистая, представляет собой круглое сооружение. Изделие, которое встречает и заполняет пространство, приветствуя посетителей и гидов. Это прелюдия к просторному внутреннему пространству, которое подчеркивает постоянные усилия компании по дизайну и совершенству.
Это прелюдия к просторному внутреннему пространству, которое подчеркивает постоянные усилия компании по дизайну и совершенству.
Однако он находится в центре офисов, в самом сердце здания, где мы находим философию компании в ее архитектурном памятнике: винтовой лестнице, спроектированной в сотрудничестве с архитектором Хосе Мария Томас Ллавадор и дизайнер и дизайнер интерьеров Хосе Косин . Как и в Actiu, эта лестница – это движение, динамизм и функциональность. Архитектурный объект без опоры, который выступает в качестве связующего звена между тремя этажами.В своей простой элегантности эта лестница выполняет три функции: соединяет пространство, декор и структуру.
Пример тщательности и внимания к деталям. Но также устойчивость, фундаментальная часть ДНК Actiu, которую мы находим в каждом из ее действий: зданиях, производственных процессах, продуктах, сырье и умах сотрудников. Так же, как технологический парк создан для обеспечения самообеспечения своими природными ресурсами, такими как вода и энергия, эта лестница была спроектирована с использованием самых инновационных производственных технологий, позволяющих разложить ее на различные стальные модули.Модули, которые были собраны и приварены на месте, , что оптимизировало их транспортировку и уменьшило количество работ.
Так же, как технологический парк создан для обеспечения самообеспечения своими природными ресурсами, такими как вода и энергия, эта лестница была спроектирована с использованием самых инновационных производственных технологий, позволяющих разложить ее на различные стальные модули.Модули, которые были собраны и приварены на месте, , что оптимизировало их транспортировку и уменьшило количество работ.
Эта лестница находится в корпоративном здании компании, одном из семи зданий, составляющих Технологический парк Actiu , экологичный промышленный проект, сертифицированный LEED Gold, присужденный Американским советом по экологическому строительству и это превратило Actiu в первую промышленную компанию в своем секторе в Европе, получившую этот сертификат.Этот технологический парк, спроектированный Хосе Мария Томас Ллавадор , иллюстрирует возможность существования модели, предназначенной для корпоративного здания, выходящей за рамки функциональности, создавая среду, в которой такие ценности, как эффективность, универсальность и экологичность, сочетаются с возвращением традиции в использовании материалов и оптимизации ресурсов. Это также парк, в котором продуманы все детали и соответствуют ценностям компании: привязанности к земле, устойчивости, важности дизайна и функциональности.
Это также парк, в котором продуманы все детали и соответствуют ценностям компании: привязанности к земле, устойчивости, важности дизайна и функциональности.
архитектурных элементов – Главная | Facebook
Компания Architectural Elements предлагает уникальный опыт изготовления. В нашей компании работают чрезвычайно квалифицированные сотрудники, которые могут предложить вам товары, которых нет больше нигде. Обладая многолетним опытом изготовления и проектирования, компания Architectural Elements раздвигает пределы возможностей изготовления металлических изделий. Мы не только создаем уникальные изделия, изготовленные по индивидуальному заказу, но и создаем душевное спокойствие в процессе. Мы хотим, чтобы вы были уверены, что ваш проект будет выполнен правильно с первого раза.Нам искренне нравится то, что мы делаем, и мы надеемся, что сможем доставить удовольствие вашему проекту. С нетерпением ждем сотрудничества с вами!– УСЛУГИ –
ЖИЛЫЕ ФАБРИКАЦИИ: Мебель для дома и на открытом воздухе, металлические столешницы, металлические двери / окна, металлические окна, стальные конструкции, водные элементы, фурнитура, решетки, металлическая облицовка, пожарные чаши и ямы, ворота, декоративные Экраны, лестницы, перила, металлические каминные двери, светильники, деревянные кронштейны
КОММЕРЧЕСКОЕ ИЗГОТОВЛЕНИЕ: скульптуры, конструкционная сталь, искусство / скульптура, металлическая облицовка, внутренняя и внешняя мебель, беседки и беседки, решетки и шпалеры, навесы и навесы , Пожарные ямы, Визуальные барьерные экраны, Водные элементы, Уличные кухни, Ворота, Скамейки, Системы перил, Лестницы, Вывески / киоски, Решетки и крышки, Удобства на объекте
ДИЗАЙН ПРОДУКТА: Анализ проекта, Ключевые требования, Исследования, Исследование концепции, Концепция Разработка, проектирование, разработка продукта, прототипирование, проектирование, тестирование, предпроизводственная разработка, производство
– ВОЗМОЖНОСТИ –
ОЦЕНКА: Наша собственная оценка partment старается, чтобы ваши сметы были выполнены вовремя. Мы предоставляем подробные и продуманные оценки, которые легко понять.
Мы предоставляем подробные и продуманные оценки, которые легко понять.
ЧЕРТЕЖ: Планы цеха – важная часть любого проекта. С тысячами часов совместной работы на САПР и на досках мы являемся экспертами в рисовании.
ИНЖИНИРИНГ: Используя штатных инженеров-технологов, мы обеспечиваем проектирование наших проектов так, чтобы они работали и функционировали так, как они задуманы.
ПРОИЗВОДСТВО: Имея полностью функциональное производственное предприятие, мы строим, тестируем и подготавливаем все в нашем магазине. Мы делаем как можно меньше аутсорсинга и стараемся производить наши изделия от “А до Я”.”
КОНТРОЛЬ КАЧЕСТВА: Качество чрезвычайно важно для нас. За последнее десятилетие мы построили нашу компанию на качестве.
УСТАНОВКА: С нашей командой по установке, состоящей из высококвалифицированных профессионалов, мы гарантируем, что ваша установка будет выполнена правильно с первого раза.
Целлюлоза как архитектурный элемент пространственно структурированных биопленок Escherichia coli
Abstract
Морфологическая форма в многоклеточных агрегатах возникает в результате взаимодействия генетической конституции и сигналов окружающей среды. Биопленки бактериальных макроколоний, которые образуют сложные трехмерные структуры, такие как большие и часто радиально ориентированные гребни, концентрические кольца и сложные морщины, предоставляют уникальную возможность понять это взаимодействие «природы и воспитания» в морфогенезе на молекулярном уровне. Морфология макроколоний зависит от компонентов внеклеточного матрикса, образующихся самостоятельно. В Escherichia coli это индуцированные стационарной фазой амилоидные курчавые волокна и целлюлоза. В то время как широко используемый «одомашненный» E.Лабораторные штаммы coli K-12 не способны генерировать целлюлозу, мы смогли восстановить производство целлюлозы и морфологию макроколоний штамма W3110 E. coli K-12 путем «ремонта» одного хромосомного SNP в опероне bcs . С помощью сканирующей электронной и флуоресцентной микроскопии целлюлозные волокна, листы и нанокомпозиты с изогнутыми волокнами были локализованы in situ при клеточном разрешении внутри физиологически двухслойных биопленок макроколоний этого «обезвреженного» штамма.
Биопленки бактериальных макроколоний, которые образуют сложные трехмерные структуры, такие как большие и часто радиально ориентированные гребни, концентрические кольца и сложные морщины, предоставляют уникальную возможность понять это взаимодействие «природы и воспитания» в морфогенезе на молекулярном уровне. Морфология макроколоний зависит от компонентов внеклеточного матрикса, образующихся самостоятельно. В Escherichia coli это индуцированные стационарной фазой амилоидные курчавые волокна и целлюлоза. В то время как широко используемый «одомашненный» E.Лабораторные штаммы coli K-12 не способны генерировать целлюлозу, мы смогли восстановить производство целлюлозы и морфологию макроколоний штамма W3110 E. coli K-12 путем «ремонта» одного хромосомного SNP в опероне bcs . С помощью сканирующей электронной и флуоресцентной микроскопии целлюлозные волокна, листы и нанокомпозиты с изогнутыми волокнами были локализованы in situ при клеточном разрешении внутри физиологически двухслойных биопленок макроколоний этого «обезвреженного» штамма. В качестве архитектурного элемента целлюлоза придает сплоченность и эластичность, т. Е. Тканеподобные свойства, которые – вместе с сеткой изогнутых волокон, покрывающих клетки, и геометрическими ограничениями в растущей колонии – объясняют образование длинных и высоких гребней и сложных морщин диких растений. тип макроколонии. Напротив, матрица биопленки, состоящая только из сети завитков волокон, является хрупкой и распадается на узор концентрических куполообразных колец, разделенных глубокими щелями. Эти исследования теперь создают основу для выяснения того, как регуляторные сети и, в частности, передача сигналов c-di-GMP работают в трехмерном пространстве высокоструктурированных и «тканеподобных» бактериальных биопленок.
В качестве архитектурного элемента целлюлоза придает сплоченность и эластичность, т. Е. Тканеподобные свойства, которые – вместе с сеткой изогнутых волокон, покрывающих клетки, и геометрическими ограничениями в растущей колонии – объясняют образование длинных и высоких гребней и сложных морщин диких растений. тип макроколонии. Напротив, матрица биопленки, состоящая только из сети завитков волокон, является хрупкой и распадается на узор концентрических куполообразных колец, разделенных глубокими щелями. Эти исследования теперь создают основу для выяснения того, как регуляторные сети и, в частности, передача сигналов c-di-GMP работают в трехмерном пространстве высокоструктурированных и «тканеподобных» бактериальных биопленок.
ВВЕДЕНИЕ
Биопленки – это многоклеточная форма бактериальной жизни, которая придает устойчивость к стрессам окружающей среды, антибиотикам и иммунной системе хозяина (1). Кроме того, сложная макроскопическая архитектура некоторых бактериальных биопленок предоставляет интригующую возможность изучить возникновение морфологических форм в многоклеточных агрегатах, обусловленных взаимодействием генетической информации и воздействий окружающей среды. Внутри биопленок экспрессия генов неоднородна, и можно наблюдать физиологически разные зоны и слои, содержащие разные компоненты внеклеточного матрикса.Эти зоны образуются в результате градиентов питательных веществ, сигнальных соединений или продуктов жизнедеятельности, которые накапливаются во время роста биопленки (2, 3). Биопленки, выращиваемые в течение нескольких дней на поверхности агара, то есть в «макроколониях», могут принимать сложные трехмерные структуры (), которые были названы «морщинистыми», «морщинистыми» или «рдарными» (для красных, сухих и шероховатых). (4–7). Понятно, что внеклеточные полимерные вещества (EPS), которые образуют матрицу, включающую адгезины, амилоид-образующие белки и экзополисахариды, необходимы для создания этих морфологических структур (8-10), но как их фактические трехмерные структуры контролируются и почему они образовались, остается неясным.
Внутри биопленок экспрессия генов неоднородна, и можно наблюдать физиологически разные зоны и слои, содержащие разные компоненты внеклеточного матрикса.Эти зоны образуются в результате градиентов питательных веществ, сигнальных соединений или продуктов жизнедеятельности, которые накапливаются во время роста биопленки (2, 3). Биопленки, выращиваемые в течение нескольких дней на поверхности агара, то есть в «макроколониях», могут принимать сложные трехмерные структуры (), которые были названы «морщинистыми», «морщинистыми» или «рдарными» (для красных, сухих и шероховатых). (4–7). Понятно, что внеклеточные полимерные вещества (EPS), которые образуют матрицу, включающую адгезины, амилоид-образующие белки и экзополисахариды, необходимы для создания этих морфологических структур (8-10), но как их фактические трехмерные структуры контролируются и почему они образовались, остается неясным.
Биосинтез целлюлозы и сложная морфология макроколоний после хромосомной «репарации» SNP, который генерирует стоп-кодон в начале bcsQ в E. coli K-12 штамм W3110. (A) Структура оперона области bcs и положение SNP TAG / TTG в bcsQ . (B) Структура макроколонии штамма AR3110, производного W3110 с аллелем bcsQ (TTG) после выращивания на бессолевых чашках LB в течение 5 дней. (C) Флуоресценция однодневных колоний, выращенных на чашках Calcofluor штамма AR3110 и его указанных мутантных производных и штамма W3110.(D) Морфология колонии AR3110 и указанных мутантных производных после роста на чашках CR в течение 5 дней.
coli K-12 штамм W3110. (A) Структура оперона области bcs и положение SNP TAG / TTG в bcsQ . (B) Структура макроколонии штамма AR3110, производного W3110 с аллелем bcsQ (TTG) после выращивания на бессолевых чашках LB в течение 5 дней. (C) Флуоресценция однодневных колоний, выращенных на чашках Calcofluor штамма AR3110 и его указанных мутантных производных и штамма W3110.(D) Морфология колонии AR3110 и указанных мутантных производных после роста на чашках CR в течение 5 дней.
В Escherichia coli и Salmonella морфология макроколонии зависит от присутствия локонных волокон и экзополисахаридной целлюлозы в матрице биопленки (7, 11, 12). Во время секреции curli белки собираются в амилоидные волокна (13, 14), которые образуют корзинообразные структуры вокруг маленьких голодающих клеток в верхнем слое макроколонии (2).Целлюлоза представляет собой полимер глюкозы с β-1,4-гликозидной связью, который у бактерий синтезируется и выводится встроенным в мембрану целлюлозосинтазным комплексом, состоящим из двух субъединиц, BcsA и BcsB (15, 16). Этот процесс поддерживается и регулируется несколькими вспомогательными белками. Участвующие белки кодируются двумя дивергентно расположенными и высококонсервативными оперонами: yhjR-bcsQABZC и bcsEFG (12, 17). И комменсальные, и патогенные штаммы E. coli гетерогенны в отношении их способности продуцировать локоны и / или целлюлозу (18, 19).
Этот процесс поддерживается и регулируется несколькими вспомогательными белками. Участвующие белки кодируются двумя дивергентно расположенными и высококонсервативными оперонами: yhjR-bcsQABZC и bcsEFG (12, 17). И комменсальные, и патогенные штаммы E. coli гетерогенны в отношении их способности продуцировать локоны и / или целлюлозу (18, 19).
Как впервые было показано для Salmonella , биосинтез curli и целлюлозы происходит в клетках ранней стационарной фазы, поскольку он зависит от регулятора биопленки CsgD, который, в свою очередь, требует для экспрессии сигма-фактора стационарной фазы RpoS (σ S ). (20, 21). Во многих штаммах E. coli CsgD регулируется по температуре и экспрессируется только ниже 30 ° C (18, 22). Производство курли и целлюлозы также регулируется повсеместно способствующим образованию биопленок вторичным мессенджером бис- (3′-5 ‘) – циклическим димерным GMP (c-di-GMP), который продуцируется и разлагается множеством дигуанилатциклаз (DGC; характеризуется GGDEF) и фосфодиэстеразы (PDE; с EAL-доменами) соответственно (23-25). c-di-GMP имеет множество компонентов для производства курли и целлюлозы. Во-первых, для экспрессии регулятора транскрипции CsgD необходимы два модуля c-di-GMP, каждый из которых состоит из DGC и PDE, действующих каскадом (26-28). Затем CsgD управляет транскрипцией генов curli ( csgBAC ), а также yaiC ( adrA в Salmonella ), который кодирует еще один DGC, необходимый для активации синтазы целлюлозы (12, 21, 28 , 29). Через c-ди-GMP-связывающий домен PilZ BcsA целлюлозосинтаза является прямой мишенью для c-ди-GMP (30).
c-di-GMP имеет множество компонентов для производства курли и целлюлозы. Во-первых, для экспрессии регулятора транскрипции CsgD необходимы два модуля c-di-GMP, каждый из которых состоит из DGC и PDE, действующих каскадом (26-28). Затем CsgD управляет транскрипцией генов curli ( csgBAC ), а также yaiC ( adrA в Salmonella ), который кодирует еще один DGC, необходимый для активации синтазы целлюлозы (12, 21, 28 , 29). Через c-ди-GMP-связывающий домен PilZ BcsA целлюлозосинтаза является прямой мишенью для c-ди-GMP (30).
Эта регуляторная сеть интенсивно изучается на молекулярном уровне. Однако вопрос о том, как этот комплексный контроль определяет пространственное распределение компонентов матрицы и, таким образом, каким-то образом трансформируется в сложную трехмерную морфологию биопленки макроколонии, был решен только совсем недавно. Макроколонии лабораторного штамма W3110 E.coli K-12 образуют узоры из концентрических колец, окруженных небольшими морщинами, и плоской областью растущих клеток на внешнем крае колонии (). Используя флуоресцентную микроскопию криосрезов, а также сканирующую электронную микроскопию (СЭМ) с высоким разрешением, маленькие клетки, плотно окруженные завитками, наблюдались в верхнем слое, тогда как палочковидные и делящиеся клетки, запутанные сеткой жгутиков, были обнаружены в верхнем слое. в нижнем слое и во внешних зонах роста макроколоний W3110. В переходных зонах между ними, пятна и небольшие цепочки из окруженных завитками и «голых» жгутиковых клеток встречаются бок о бок, что также отражается в гетерогенной экспрессии CsgD (2).Это исследование было первым, которое продемонстрировало физиологическую дифференциацию в биопленке макроколонии in situ при клеточном разрешении и тем самым прояснило, что волокна curli и RpoS-зависимая экспрессия генов ( csgD и генов curli) происходят в верхнем слое и на поверхности. .
Используя флуоресцентную микроскопию криосрезов, а также сканирующую электронную микроскопию (СЭМ) с высоким разрешением, маленькие клетки, плотно окруженные завитками, наблюдались в верхнем слое, тогда как палочковидные и делящиеся клетки, запутанные сеткой жгутиков, были обнаружены в верхнем слое. в нижнем слое и во внешних зонах роста макроколоний W3110. В переходных зонах между ними, пятна и небольшие цепочки из окруженных завитками и «голых» жгутиковых клеток встречаются бок о бок, что также отражается в гетерогенной экспрессии CsgD (2).Это исследование было первым, которое продемонстрировало физиологическую дифференциацию в биопленке макроколонии in situ при клеточном разрешении и тем самым прояснило, что волокна curli и RpoS-зависимая экспрессия генов ( csgD и генов curli) происходят в верхнем слое и на поверхности. .
Однако целлюлоза не могла быть обнаружена в этом исследовании, поскольку штаммы E. coli K-12 не производят ее (12, 19). Причина этого дефицита неизвестна, начиная с г. coli K-12 действительно имеет опероны bcs и экспрессирует все регуляторные гены, включая csgD и ген DGC yaiC (27, 28). В настоящем исследовании мы проследили целлюлозно-отрицательный фенотип до «одомашнивания» однонуклеотидной мутации. «Ремонт» этого SNP в хромосоме позволил нам ответить на ряд вопросов в хорошо охарактеризованном штамме E. coli K-12, например, где в биопленке мы находим целлюлозу? Как выглядит его микроструктура на клеточном уровне, в частности, при контакте с другими компонентами матрикса, такими как волокна курчавых и жгутиков? Как целлюлоза способствует формированию трехмерной структуры и физическим свойствам биопленок макроколоний? Наши данные демонстрируют, что целлюлоза является архитектурным элементом с четким пространственным распределением внутри биопленки.Вместе с амилоидными локонами он образует нанокомпозитный «строительный материал», который придает тканеподобные свойства, то есть сцепление, эластичность и стабильность, которые в сочетании с пространственными ограничениями в растущих колониях создают замечательные элементы трехмерной структуры.
coli K-12 действительно имеет опероны bcs и экспрессирует все регуляторные гены, включая csgD и ген DGC yaiC (27, 28). В настоящем исследовании мы проследили целлюлозно-отрицательный фенотип до «одомашнивания» однонуклеотидной мутации. «Ремонт» этого SNP в хромосоме позволил нам ответить на ряд вопросов в хорошо охарактеризованном штамме E. coli K-12, например, где в биопленке мы находим целлюлозу? Как выглядит его микроструктура на клеточном уровне, в частности, при контакте с другими компонентами матрикса, такими как волокна курчавых и жгутиков? Как целлюлоза способствует формированию трехмерной структуры и физическим свойствам биопленок макроколоний? Наши данные демонстрируют, что целлюлоза является архитектурным элементом с четким пространственным распределением внутри биопленки.Вместе с амилоидными локонами он образует нанокомпозитный «строительный материал», который придает тканеподобные свойства, то есть сцепление, эластичность и стабильность, которые в сочетании с пространственными ограничениями в растущих колониях создают замечательные элементы трехмерной структуры. биопленок макроколоний.
биопленок макроколоний.
МАТЕРИАЛЫ И МЕТОДЫ
Штаммы бактерий.
Все использованные штаммы являются производными штамма E. coli K-12 W3110 (31). Мутации rpoS :: cat , csgD :: cat , csgB :: cat и bcsA :: cat представляют собой кассету с делецией (Δ) / резистентностью в полностью открытой рамке считывания вставки, описанные ранее (2, 27, 28).Для создания мутантов с делецией / вставкой bcsQ :: cat , bcsEFG :: kan и yhjR :: cat с делецией / вставкой был применен протокол одностадийной инактивации (32) с использованием олигонуклеотидных праймеров. перечислены в Таблице S1 в дополнительном материале. При необходимости кассеты перекидывались (32). Эти мутации были перенесены путем трансдукции P1 (33).
Для замены кодона 6 bcsQ с TAG (стоп) на TTG (лейцин) в хромосомной копии bcsQ в штамме W3110, двухэтапная процедура замены, связанная с протоколом одноэтапной инактивации (32) был применен. Олигонуклеотиды для этой процедуры перечислены в таблице S1 дополнительного материала. В процедуре используется фрагмент плазмиды pKD45 (32), кодирующий кассету устойчивости к канамицину и токсин ccdB под контролем индуцируемого рамнозой промотора, который сначала вводится в целевой локус хромосомы с последующей его заменой. путем рекомбинации с желаемым аллелем (на фрагменте ПЦР, преобразованном в реципиента), который выбирается путем роста в присутствии рамнозы (34).Аллельные состояния трансформантов проверяли с помощью ПЦР и секвенирования ДНК.
Олигонуклеотиды для этой процедуры перечислены в таблице S1 дополнительного материала. В процедуре используется фрагмент плазмиды pKD45 (32), кодирующий кассету устойчивости к канамицину и токсин ccdB под контролем индуцируемого рамнозой промотора, который сначала вводится в целевой локус хромосомы с последующей его заменой. путем рекомбинации с желаемым аллелем (на фрагменте ПЦР, преобразованном в реципиента), который выбирается путем роста в присутствии рамнозы (34).Аллельные состояния трансформантов проверяли с помощью ПЦР и секвенирования ДНК.
Построение единственной копии
lacZ репортерных сплавов. Олигонуклеотидные праймеры, используемые для конструирования слитых однокопийных репортерных генов lacZ слитых yhjR , bcsQ и bcsA , перечислены в таблице S1 в дополнительном материале. Процедура с использованием вектора слияния lacZ pJL28 была описана ранее (28). Все слияния являются трансляционными слияниями, которые включают 9 ( yhjR ), 18 ( bcsQ ; отсчет от стартового кодона ATG;) и 10 ( bcsA ) кодонов соответствующих открытых рамок считывания. Вышестоящие области всех трех слитых репортерных генов lacZ содержат всю межгенную область между оперонами yhjR-bcsQABZC и bcsEFG и начинаются на 406 п.н. перед стартовым кодоном yhjR (см. Также рис. S1 в дополнительный материал). Все репортерные слияния переносили в положение att (λ) хромосомы производного W3110, содержащего делецию Δ lac ( I-A ) :: шрам , через фаг λRS45 или λRS74 (35).Единичную лизогению тестировали методом ПЦР (36).
Вышестоящие области всех трех слитых репортерных генов lacZ содержат всю межгенную область между оперонами yhjR-bcsQABZC и bcsEFG и начинаются на 406 п.н. перед стартовым кодоном yhjR (см. Также рис. S1 в дополнительный материал). Все репортерные слияния переносили в положение att (λ) хромосомы производного W3110, содержащего делецию Δ lac ( I-A ) :: шрам , через фаг λRS45 или λRS74 (35).Единичную лизогению тестировали методом ПЦР (36).
Экспрессия слияния репортера lacZ с yhjR , bcsQ и bcsA в присутствии или отсутствии E. coli K-12-специфического стоп-кодона в bcsQ . Производные штамма W3110, несущие единственную копию слияния lacZ в yhjR или в bcsQ или bcsA с кодоном 6 в bcsQ , являющимся либо TAG (слияния W), либо TTG (слияния AR), выращивали в среде LB при 28 ° С.Определяли оптическую плотность (светлые символы) и удельную активность β-галактозидазы (темные символы).
Генерация хромосомных аллелей, кодирующих 3 × FLAG-меченых белка.
Хромосомно кодируемых конструкций с 3 × FLAG-тегами на С-конце были созданы с использованием плазмиды pSUB11 в качестве матрицы ПЦР (37) и олигонуклеотидных праймеров, перечисленных в таблице S1 в дополнительном материале, в соответствии с процедурой, основанной на технологии λRed (32).
Рост бактериальных макроколоний.
Клетки выращивали в течение ночи в жидкой среде Лурия-Бертани (LB) (33) при аэрации при 37 ° C. Всего 5 мкл ночных культур наносили на чашки с бессолевым агаром LB. Где указано, в чашки с агаром добавляли конго красный (CR; 40 мкг мл -1 ) и бриллиантовый синий Кумасси (20 мкг мл -1 ; чашки CR), Calcofluor (100 мкг мл -1 ) или Тиофлавин S (TS; 40 мкг мл -1 ). Для параллельного выращивания всех сравниваемых штаммов на одной чашке с агаром были использованы чашки Петри диаметром 140 мм (VWR), что позволяет выделить до 25 макроколоний в матрице 5 × 5 на одной чашке.Чашки с макроколониями инкубировали при 28 ° C до 7 дней. Целлюлозу определяли по флуоресценции колоний на планшете Calcofluor под УФ-светом 366 нм.
Стереомикроскопия.
Биопленки макроколоний E. coli визуализировали при увеличении × 10 с помощью стереомикроскопа Stemi 2000-C (Zeiss, Oberkochen, Германия). Цифровые изображения были получены с помощью цифровой камеры AxioCamICC3, соединенной со стереомикроскопом, управляемой с помощью программного обеспечения AxioVision 4.8 (Zeiss).
Запись видео.
Макроколонии E. coli на чашках с агаром были полностью покрыты культуральной средой и подвергались легкому встряхиванию. Фильмы растворения макроколоний под действием сил сдвига регистрировали в реальном времени с помощью камеры Cyber-Shot DSC (Sony, Токио, Япония).
Криосрезы биопленок макроколоний и флуоресцентная микроскопия.
Процедура и материалы, использованные для криосрезов биопленок макроколоний и дальнейшего изучения и визуализации флуоресценции тиофлавина S в поперечных срезах, были точно такими, как описано ранее (2).
СЭМ бактериальных макроколоний.
Подготовка и последующий анализ образцов биопленок макроколоний с помощью СЭМ выполняли точно так же, как описано ранее (2).
Определение активности β-галактозидазы.
Активность β-галактозидазы оценивалась с использованием –-нитрофенил-β-d-галактопиранозида (ONPG) в качестве субстрата и выражалась в мкмоль из – -нитрофенола в минуту на мг клеточного белка (33). . Эксперименты, показывающие экспрессию слияний lacZ на протяжении всего цикла роста, были выполнены, по крайней мере, дважды, и показаны результаты репрезентативного эксперимента.
РЕЗУЛЬТАТЫ
Восстановление биосинтеза целлюлозы у «одомашненных»
E. coli K-12 коренным образом изменяет морфологию и физические свойства биопленок макроколоний.В поисках предполагаемых мутаций, которые могут объяснить целлюлозно-отрицательный фенотип E. coli K-12, мы сравнили опероны bcs в геномных последовательностях штаммов E. coli K-12 W3110 и MG1655 с соответствующими геномными участками синтезирующего целлюлозу E.coli , в частности, энтероагрегантный (EAEC) штамм 55989, который не только продуцирует целлюлозу, но и развивается в структурно сложные макроколонии (19). В штаммах E. coli K-12 кодон GTG был аннотирован как стартовый кодон гена bcsQ (). Однако мы наблюдали предполагаемый стоп-кодон (TAG) всего на три кодона выше по течению с другим предполагаемым стартовым кодоном, расположенным на пять кодонов выше по течению, что не только является оптимальным ATG, но также предшествует, казалось бы, лучшая последовательность Шайна-Далгарно, чем стартовый кодон GTG. изначально аннотировано.Это предполагает, что трансляция bcsQ может фактически начинаться с кодона ATG, но может быть завершена только пятью кодонами ниже по течению, что может привести к полярности экспрессии генов ниже по течению от bcsQ . Более того, продуцирующий целлюлозу штамм 55989 (а также многие другие штаммы E. coli разного происхождения и свойств) несут кодон TTG вместо этого предполагаемого стоп-кодона TAG в bcsQ . Кроме того, мы наблюдали еще несколько однонуклеотидных полиморфизмов (SNP) в bcs оперонах E.coli K-12 и EAEC 55989, но эти SNP приводят либо к синонимичным кодонам, либо к консервативным заменам аминокислот.
Таким образом, SNP TAG / TTG на ранней стадии bcsQ явно имел наибольшие потенциальные последствия для экспрессии гена bcs . Поэтому мы заменили ТАГ в хромосоме штамма W3110 на TTG, используя двухэтапную процедуру замены, основанную на технологии λRed. «Ремонт» этого SNP привел к резкому изменению морфологии колонии. Макроколонии полученного штамма AR3110 были очень большими (с диаметром> 20 мм через 5-7 дней), были чрезвычайно плоскими, с радиальными гребнями и сложными морщинами, выступающими из плоских участков колоний ().Эти макроколонии окрашивались в темно-красный цвет амилоидным красителем Конго красным (CR), но морфологические структуры также формировались на чашках с агаром без CR. Количество гребней и морщин увеличивалось на относительно влажных пластинах с агаром, создавая узоры, напоминающие морфотип «rdar» (), первоначально описанный для Salmonella (7), а также наблюдаемый для некоторых патогенных E. coli (18, 38), тогда как макроколонии, выращенные на несколько более сухих пластинах агара, образовывали меньше радиальных гребней высотой до 5 мм ().Эти радиальные гребни начали формироваться во внешних областях макроколоний между одним и двумя днями роста, при этом тонкий узор радиальных линий был виден уже в первый день, некоторые из которых выгибались в гребни, тогда как промежуточные, казалось, сглаживались. из. Затем гребни медленно увеличивались в высоту и распространялись к центральной части макроколонии ().
Формирование архитектуры макроколонии с течением времени. На центральных панелях показаны изображения с низким увеличением макроколоний W3110 и AR3110, развивающихся с течением времени на планшетах LB, содержащих TS, не содержащих соли.Верхняя и нижняя панели показывают увеличенные изображения макроколоний, представленных на центральных панелях.
Связывание и флуоресценция Calcofluor показали, что макроколонии AR3110 генерируют целлюлозу (). Более того, точная делеция гена целлюлозосинтазы bcsA в AR3110 воссоздала морфологию кольцевой структуры, ранее наблюдаемую для штамма W3110 (). Сходным образом точные и неполярные делеции bcsQ и yhjR и – в меньшей степени – делеция оперона bcsEFG в AR3110 приводили к образованию макроколоний с кольцевыми структурами, указывающими на отсутствие или снижение биосинтеза целлюлозы.Производное, удаленное csgB , которое синтезирует целлюлозу, но не содержит curli-волокон, вызывало розовое окрашивание пластин CR и узор из плотных мелких морщинок ().
Мы пришли к выводу, что SNP, который генерирует стоп-кодон в начале bcsQ , вмешивается в биосинтез целлюлозы в «одомашненных» лабораторных штаммах E. coli K-12. Чтобы количественно оценить этот эффект в отношении экспрессии генов bcs ниже этого стоп-кодона, мы сгенерировали однокопийные слияния lacZ в yhjR , bcsQ и bcsA (см.рис.S1 в дополнительном материале), причем слияния последних двух генов содержат либо стоп-кодон TAG, как в W3110 (‘ WbcsQ :: lacZ ‘ и ‘ WbcsA :: lacZ ‘), либо TTG кодон, присутствующий в AR3110 (‘ ARbcsQ :: lacZ ‘ и ‘ ARbcsA :: lacZ ‘). Экспрессия репортерных слияний с кодоном TTG была действительно более чем в 10 раз выше, чем экспрессия слияния, несущего стоп-кодон в этом положении (), независимо от того, анализировалась ли экспрессия слияния в отсутствие или в присутствии продукции целлюлозы (i .е., в генетическом фоне W3110 или AR3110). Данные слияния lacZ также показывают, что экспрессия yhjR , т.е. первого гена в опероне, функция которого в биосинтезе целлюлозы неизвестна, была в ~ 5 раз выше, чем экспрессия bcsQ и bcsA . Более того, все три гена (и, следовательно, вероятно, весь оперон) уже показали некоторую экспрессию в растущих клетках, которая далее увеличивалась в зависимости от σ S , когда клетки вступали в стационарную фазу ().Используя маркировку флагом bcsA в хромосоме и иммуноблоттинг, мы могли также обнаружить целлюлозосинтазу в макроколониях репарированного штамма AR3110, но не в W3110 (см. Рис. S2 в дополнительном материале).
Локализация целлюлозных и амилоидных волокон курли в
макроколониях E. coli .Восстановление биосинтеза целлюлозы в штамме AR3110 позволило нам изучить функцию и точную локализацию целлюлозы в матрице биопленок макроколоний этого хорошо охарактеризованного в остальном E.coli K-12. Макроколонии выращивали на бессолевой среде LB с добавлением тиофлавина S (TS), флуоресцентного красителя, который, как и CR, связывает амилоидные волокна локонов, а также целлюлозу без изменения структуры макроколоний (). Макроколонии анализировали с помощью криосрезов в сочетании с светлопольной и флуоресцентной микроскопией.
В соответствии с макроскопической морфологией колоний, уже при малом увеличении исследование в светлом поле поперечных срезов зрелых макроколоний AR3110 выявило драматические архитектурные изменения, связанные с восстановленной экспрессией оперона bcs и производством целлюлозы.Плоские участки в центре макроколонии или между гребнями имели очень правильную высоту только ~ 60 мкм, которые, тем не менее, дифференцировались на два отдельных слоя (). Верхний бактериальный слой (толщиной около 40-50 мкм) демонстрировал интенсивную флуоресценцию TS, тогда как нижний слой клеток (толщиной ~ 20 мкм) не имел флуоресценции. Флуоресценция TS специфична для curli и целлюлозы, поскольку она была обнаружена, когда присутствовали только curli или целлюлоза (и), но полностью отсутствовала у мутанта, который не продуцировал ни один из двух компонентов матрицы ().В очень плоской зоне роста на внешнем крае макроколоний AR3110 (см. Рис. S3 в дополнительном материале) флуоресценция TS не могла быть обнаружена, тогда как на глубине> 200 мкм внутри начиналась нарастать двухслойная структура.
Ограниченная локализация curli и целлюлозы определяет двухслойную архитектуру макроколоний E. coli . Пятидневные биопленки макроколонии E. coli , выращенные на бессолевой среде LB с добавлением TS, криозаждали и делали срезы перпендикулярно плоскости макроколонии толщиной 5 мкм.Тонкие срезы визуализировали при малом увеличении, с яркими полями (левые панели), флуоресцентными (средние панели) и объединенными (правые панели) микроскопическими изображениями, представляющими поперечные сечения макроколоний W3110 (A), AR3110 (B) и его Δ bcsA (C), Δ csgB (D) и Δ csgB / Δ bcsA (E) производные. Флуоресцентные изображения были ложно окрашены в зеленый цвет для TS. Изображения визуализируют весь вертикальный разрез макроколоний. Верхние левые вставки в изображениях в светлом поле показывают изображения соответствующих макроколоний в виде сверху.
Образование вертикального гребня начинается с выпучивания во внешней зоне плоской колонии, в результате чего образуются двойные слои, которые затем перемещаются вверх и поддерживают постоянную ширину независимо от фактической высоты гребня. Формирование гребня зависело как от локонов, так и от целлюлозы. Макроколонии штаммов, которые производят curli, демонстрируют только концентрические кольца (), которые в поперечных сечениях выглядят как серия смежных куполообразных возвышений> 200 мкм высотой (и). Вместо этого, когда целлюлоза производилась в отсутствие завитков, макроколония показывала узор из небольших переплетенных морщин в центральной зоне колонии ().В поперечных сечениях это превратилось в волнистую поверхность (). Отсутствие как локонов, так и целлюлозы привело к образованию толстой (~ 200 мкм высотой) макроколонии без какой-либо морфологической дифференциации ().
Вертикальные гребни представляют собой узкий внутренний слой нефлуоресцентных ячеек (). Примечательно, что бактерии в этом внутреннем слое не включали образование локонов / целлюлозы, даже когда они были расположены близко к вершине гребня, то есть на большом расстоянии от агара, обеспечивающего питательные вещества. Следует также отметить, что пространство у основания гребней, которое открывается во время коробления макроколонии в этом положении, было полностью заполнено нефлуоресцентными клетками, предполагая, что это пространство было заполнено растущими клетками.
Чтобы узнать о подробном пространственном расположении целлюлозы в макроколониях, мы сначала внимательно изучили штамм, который продуцирует только целлюлозу (AR3110 Δ csgB ). Здесь целлюлоза была визуализирована по картине флуоресценции TS, которая повторяла форму мелких морщинок. Вогнутые области, образовавшиеся ниже и выше волнистого целлюлозосодержащего клеточного слоя, также оказались занятыми бактериями (). Дальнейшее увеличение флуоресцентного слоя в разных местах (чтобы) выявило регулярный узор из пятен и особенно ярких струн, т.е.например, нити и, возможно, листовые структуры (длиной от 2 до 15 мкм), которые были ориентированы в основном параллельно друг другу, но перпендикулярно линии морщин. В местах, где флуоресцентный слой формировал поверхность колонии, также были обнаружены струны лежащие на поверхности (см. Стрелку на). Эти результаты не только показывают целлюлозу как компонент нитевидной матрицы, что согласуется с ее фибриллярной природой, но и демонстрируют, что ее распределение и расположение не случайны, а локализованы и высоко упорядочены.В отличие от curli-волокон, которые образуют сеть, похожую на «соты», плотно окружающие клетки, волокна целлюлозы, по-видимому, проходят между клетками и вдоль них.
Целлюлоза и curli обнаруживают дифференциальное распределение и специфическое пространственное расположение в макроколониях E. coli . (A) Изображение с малым увеличением, показывающее флуоресценцию TS в репрезентативном поперечном сечении макроколонии AR3110 csgB , как показано на. (B – D) Увеличенные изображения картины флуоресценции TS в соответствующих областях макроколоний с цветовой кодировкой, заключенных в рамку A.Стрелка в D указывает на целлюлозную нить, лежащую на поверхности. (E) Изображение с малым увеличением, показывающее флуоресценцию TS в репрезентативном поперечном сечении макроколонии AR3110, как представлено на. (F до H) Увеличенные виды картины флуоресценции TS в соответствующих областях макроколоний с цветовой кодировкой, заключенных в рамку на панели E. На вставках на панелях F и G показаны увеличенные виды соответствующих цветных областей.
Проанализировав распределение волокон curli и целлюлозы, каждое «по отдельности», мы внимательно изучили картину флуоресценции TS в макроколониях AR3110, которые продуцируют как волокна curli, так и целлюлозу.Были исследованы три репрезентативных области макроколонии: плоская область, близкая к поверхности агара, а также средняя часть и вершина гребня (). В верхнем флуоресцентном слое плоского участка четко выделялись два паттерна флуоресценции ТС (). В верхней зоне виден «пчелиный» узор вокруг силуэтов бактерий. Далее внутри верхнего слоя, то есть ближе к границе раздела с нефлуоресцентным нижним слоем, эти силуэты бактерий казались более выстроенными и вертикально ориентированными, с интенсивно флуоресцентными нитями, которые напоминают волокна целлюлозы при отсутствии изгибов (к) – возможно, образуя оболочковидные структуры (см. увеличенный вид областей, выделенных белыми рамками).Эти рисунки предполагают, что сетка из локонных волокон относительно равномерно распределена в верхнем слое, тогда как основная масса целлюлозы будет преобладать в несколько более низкой зоне верхнего слоя, то есть ближе к границе раздела с нижним слоем.
Было обнаружено, что этот узор стабильно сохраняется в средней части гребня () и был еще более выраженным, с еще более длинными интенсивно флуоресцентными волокнами (область в рамке) на кончике, где весь слой сильно изогнут (ср. а также ).Вытянутый по вертикали вид волокон и предполагаемых оболочек, а также растянутый по горизонтали вид сети изогнутых волокон вблизи поверхности, вероятно, отражает напряжение, которое вызывает крайняя кривизна на конце. Поразительное наблюдение, что поверхность зон кончика не разрушается под этим натяжением и что гребни на самом деле могут становиться более чем в 30 раз выше своей ширины без потери своей сложной пространственной архитектуры, демонстрирует, что целлюлоза придает сильное сцепление и эластичность верхнему слою макроколония.В целом, эти открытия впервые выявили сильно дифференцированно упорядоченное пространственное расположение целлюлозных волокон и сетей изогнутых волокон и предполагают, что комбинация этих двух компонентов матрикса придает тканеподобные свойства биопленкам макроколонии E. coli .
Тонкая структура целлюлозных волокон и структур из вьющихся целлюлозных композитов при клеточном разрешении в различных зонах макроколоний
E. coli .СЭМ с высоким разрешением позволил нам изучить эту пространственную архитектуру и внешний вид целлюлозы и завитков in situ на клеточном уровне.СЭМ-изображения срезов через небольшой гребень AR3110 () и морщинистую область макроколонии, состоящей только из целлюлозы (AR3110 Δ csgB ;), демонстрируют превосходное соответствие морфологическим формам, видимым на криосрезах (и). При более высоком разрешении мы сосредоточились не только на наличии и пространственном расположении целлюлозы и завитков, но и на отличительных признаках физиологического состояния бактерий, таких как размер и форма клеток, деление клеток и образование жгутиков.
Комбинированный материал из целлюлозы и вьющихся волокон покрывает поверхность макроколонии AR3110, а жгутики образуют плотную сетку внизу.(A) СЭМ-изображение в перспективе с малым увеличением макроколонии AR3110, на котором виден небольшой гребень. (B) СЭМ-изображение в перспективе с малым увеличением макроколонии Δ csgB , производной AR3110, с небольшими переплетенными морщинами. (C – E) СЭМ-изображения с высоким разрешением (увеличение 50000x), показывающие поверхности макроколоний, образованных штаммом AR3110 и его указанными производными. (F) СЭМ-изображение с высоким разрешением (увеличение × 50000), показывающее детали сетки перепутанных нитей, образованных бактериями в нижней части макроколонии AR3110, которая отсутствует в макроколонии AR3110 Δ fliC , демонстрирующая удлиненный стержень без жгутиков. -образные клетки, расположенные в плоскости (G).(H) СЭМ-изображение с высоким разрешением (увеличение 50 000 ×), показывающее длинные листы целлюлозы, взаимодействующие со жгутиками в средней внутренней зоне макроколонии AR3110 Δ csgB .
СЭМ-изображение с большим увеличением поверхности макроколонии Δ bcsA () показало ту же структуру только завитков, о которой ранее сообщалось для макроколоний W3110 (2), т.е. ). Напротив, бактериальные клетки на поверхности макроколонии Δ csgB , состоящей только из целлюлозы, не были покрыты оболочкой, а лишь частично покрыты плоскими листами неправильной формы ().Этот пластинчатый матричный материал представлял собой конкретно целлюлозу, поскольку выбивание bcsA в уже имеющем отрицательную локонность Δ csgB мутантном фоне привело к тому, что бактерии на поверхности полностью лишились матричного материала (данные не показаны).
Напротив, поверхность макроколонии AR3110 была почти полностью покрыта огромным количеством матричного материала, так что бактерии больше не были видны (). Клетки, казалось, были покрыты гладким слоем, а также соединены нитями, но изогнутые волокна и целлюлозу больше нельзя было различить, что позволяет предположить, что вместе они образуют композитный материал.Для всех исследуемых штаммов очень маленькие и яйцевидные клетки были обнаружены на поверхности макроколонии (to), что указывает на то, что клетки находятся в стационарной фазе, то есть в состоянии, при котором продуцируются оба компонента матрикса. Этот поверхностный узор регулярно наблюдался на всех участках поверхности макроколонии (например, в плоских секторах, гребнях), которые на макроуровне демонстрировали окрашивание CR.
СЭМ-исследование касающейся агара нижней зоны макроколонии AR3110 показало ту же картину (), которая наблюдалась ранее в макроколониях W3110 (2), т.е.е., плотная сетка, образованная длинными и запутанными жгутиками, отсутствующая у мутанта Δ fliC (). Эти закономерности не зависели от curli и целлюлозы, поскольку они также наблюдались в нижней части макроколоний Δ csgB , Δ bcsA и Δ csgB Δ bcsA , производных AR3110 (данные не показаны). Однако внутри макроколоний также могут быть обнаружены связи между целлюлозой и жгутиками, при этом сеть запутанных жгутиков, по-видимому, служит каркасом для образования целлюлозных нитей и удлиненных листов ().
Эти внутренние зоны биопленки были исследованы с помощью SEM в четырех положениях макроколоний AR3110: вершине, средней части и основании гребня и на плоских участках (). Во всех этих положениях области, близкие к поверхности колонии, были очень компактными с небольшими яйцевидными клетками, окруженными корзинообразными структурами, типичными для сетей curli-волокон (красные стрелки и). Далее внутри во всех четырех положениях этот образец изменился с небольшими бактериями, только частично завернутыми в длинные листы и оболочки (синие стрелки и), которые коррелируют с удлиненными структурами, визуализированными TS (to).Далее внутри наблюдались целые листы фибриллярного материала (синяя стрелка), что коррелирует с сильной флуоресценцией TS в этой области (to). Эта явно богатая целлюлозой зона очерчивала границу с самым внутренним клеточным слоем в гребнях и с нижним слоем на плоских участках. В поперечных сечениях плоской поверхности эти верхний и нижний слои также легко разрушались во время подготовки (и). Разделение на два физиологически различных типа клеток было особенно заметно в основании гребней, где небольшие клетки с неподвижной фазой, заключенные в матрицу, можно увидеть над линией деления, тогда как на нижней стороне длинные палочковидные клетки росли вертикальными рядами и заполнили пространство у основания конька (к).
Пространственное расположение целлюлозы, завитков и жгутиков и физиология клеток в контексте архитектуры двухслойной биопленки. Для обзора в левом столбце изображений показаны СЭМ-изображения с малым увеличением поперечных сечений AR3110 на вершине (A), среднем теле (D) и основании (G) гребня и плоской области (J). . В среднем и правом столбцах показаны изображения с цветными рамками с увеличением × 12000, соответствующими зонам с цветными рамками на обзорных изображениях. Красная стрелка на панели B указывает на структуру матрицы в виде корзины, типичную для изогнутых волоконно-оптических сетей.Синие стрелки на панелях C, E, F и H обозначают листовой материал, указывающий на целлюлозу. Желтая стрелка на панели E указывает на похожие на жгутики филаменты во внутренней области гребня, характеризующиеся удлиненными растущими клетками. Желтые стрелки на панелях H и I указывают на палочковидные бактерии в зоне роста ниже гребня. Красная стрелка на панели K указывает заключенные в матрицу бактерии в верхнем слое с голодающими клетками. Синие и желтые стрелки на панели L указывают клетки, контактирующие с пластинчатым и нитевидным матричным материалом в основании верхнего слоя колоний, и удлиненные палочковидные клетки, запутанные жгутиками в нижнем слое, соответственно.
Целлюлоза вместе с волокнами curli создает сцепление и эластичность и тем самым определяет морфологические формы биопленок макроколонии
E. coli .Колонии целлюлозо-положительного штамма AR3110 проявляли более сильное сцепление, например, во время повторного разделения, чем колонии, образованные родительским штаммом W3110, фенотип, также связанный с морфотипом колонии rdar в Salmonella (20, 21). Эту разницу можно легко визуализировать, покрыв полностью выросшие макроколонии жидкостью, а затем подвергнув их усилию сдвига, слегка встряхнув чашки с агаром вручную.Мутант с дефицитом курли и целлюлозы просто выпустил клеточную дымку, растворяющуюся в суспензии во время этой процедуры (см. Movie S1 в дополнительном материале). Макроколония W3110 – с локонами в качестве единственного компонента матрицы в ее верхнем слое – распалась на жесткие части, которые уплыли и распались дальше (см. Фильм S2 в дополнительном материале). Напротив, макроколония AR3110 с ее композитной матрицей из локонных волокон и целлюлозы вела себя больше как ткань (см. Фильм S3 в дополнительном материале).Таким образом, его толстый верхний слой, отделившийся от агара, поддерживает, по существу, как целую макроколонию, с его краями, «колеблющимися» взад и вперед с потоком жидкости, тогда как нижний слой, который отражает структуру морщин отделенного верхнего слоя, остался на агар. Такое поведение указывает на то, что целлюлоза в сочетании с вьющимися волокнами придает биопленке макроколонии не только сильную когезию, но и высокую эластичность – свойство, согласующееся с почти полным отсутствием разрывов на изображениях с высоким разрешением поверхности макроколоний AR3110 или ее curli-дефицитное производное целлюлозы (и).Эта выраженная эластичность, вероятно, является предпосылкой для образования сильно искривленных поверхностей, связанных с гребнями и морщинами макроколоний.
С другой стороны, жесткость и хрупкость белковой матрицы, состоящей только из сети curli-волокон, кажется основой для образования макроскопических кольцевых структур в макроколониях W3110 и других штаммов curli + целлюлозы – генерируется в настоящем исследовании (). Формирование этих колец, по-видимому, началось с поверхностных разрывов в круговой зоне растущей макроколонии, где слой скрученных клеток был еще относительно тонким и менее конфлюэнтным.Эти большие разрывы, которые, вероятно, возникли из-за давления, создаваемого все еще распространяющимися ячейками внизу, расширились в канавки (), при этом соседние изогнутые слои ячеек округлились вверх и, наконец, приняли куполообразную форму в поперечных сечениях через кольцевую структуру зрелая макроколония (). Во время этого процесса бороздки далее углублялись в щели (и), которые уходили глубоко в колонии () и которые впоследствии были колонизированы клетками, которые, казалось, выталкивались вверх из нижних слоев ().Первоначально эти клетки возникли как палочковидные жгутиковые клетки (), но, вероятно, из-за голодания на своем новом месте, позже они включили производство локонов (), что в конечном итоге могло закрыть отверстие в структуре макроколонии «вторичной» сеткой локонов. слой. Кроме того, небольшие микротрещины также образовывались на участках, покрытых сливными завитками (и к). Эти крошечные трещины, казалось, медленно открывались, и иногда поперек них тянулись волокна завитков, что не влияло на макроскопическую структуру колонии в целом.
Глубокие разрывы на поверхности макроколоний штаммов, которые производят завитки, но не целлюлозу, создают морфологию концентрических колец. (A) Изображение вида сверху зрелой макроколонии W3110, выращенной при 28 ° C на пластине CR, показывающее завиток-зависимый кольцевой узор. (B) Флуоресцентное изображение при малом увеличении поперечного сечения макроколонии W3110, выращенной в присутствии TS. (C) Увеличенное изображение колец макроколонии W3110, вид сверху. Стрелка указывает на естественные разрывы в верхней части колец.(D, E и F) SEM-изображения, показывающие изменения на поверхности макроколонии W3110 в процессе формирования кольца. Стрелка на панели F указывает бактерии, колонизирующие расщелину в более поздние моменты времени. (G) СЭМ-изображение поверхности колонии (при увеличении × 24000), показывающее начальных бактериальных колонизаторов расщелины (день 6, и далее, на панели F), которые имеют форму палочки и несут жгутики. (H) СЭМ-изображение (при увеличении × 12000), показывающее бактерии позже во время колонизации щели (день 7, и далее), которые почти заполнили щель, стали короче и частично начали генерировать завитки вокруг своих клеточных тел. .(I, J и K) СЭМ-изображения, показывающие микротрещины в верхней части зрелой макроколонии W3110 (при увеличениях × 6000, × 12000 и × 24000 соответственно).
В заключение, волокна curli и целлюлоза по отдельности или в комбинации определяют, какие трехмерные структурные элементы возникают в биопленке макроколонии. Концентрические кольца возникают как разрывы в хрупких макроколониях, которые в больших количествах образуют исключительно завитые волокна, тогда как образование гребней и сложных морщин начинается с изгиба эластичных тканеподобных макроколоний, содержащих целлюлозу, с дальнейшим подъемом устойчивых гребней, требующих завитков / матрица целлюлозного композита.
ОБСУЖДЕНИЕ
Биопленки макроколонии E. coli демонстрируют двухслойную архитектуру, которая отражает принципиально разные физиологические состояния.Ранее мы показали, что дефицитные по целлюлозе клетки E. coli K-12 (штамм W3110) образуют биопленки макроколонии, состоящие из двух физиологически различных слоев клеток (2): растущие жгутиковые клетки на внешних краях колонии, а также нижний слой, в отличие от небольших ячеек с неподвижной фазой, окруженных толстой сеткой изогнутых волокон в верхнем слое и на поверхности (кроме внешних краев).Несмотря на радикально измененную макроскопическую морфологию, «отремонтированный» целлюлозой штамм AR3110 в равной степени демонстрирует микроскопическую двухслойную архитектуру (). Из-за плоской поверхности колоний переход между этими слоями еще более резкий, с тонким нижним слоем растущих клеток – высотой всего ∼20 мкм, образующим плотную сетку жгутиков, неотличимых от таковой у не содержащего целлюлозы W3110 (; сравните с изображениями, показанными в ссылке 2). Верхний слой, который имеет высоту от 40 до 50 мкм, теперь производит волокна завитков, а также целлюлозу (, и; см.также рис.S3 в дополнительном материале), с целлюлозой, образующей толстые нити и оболочковидные структуры, которые проходят вертикально в верхний слой, а также толстые и плоские листы, которые очерчивают границу между двумя слоями, в частности, внутри гребней ().
Важно отметить, что эта двухуровневая архитектура не является следствием формирования матрицы. Скорее, это функция градиента питательных веществ, которые поступают из твердой питательной среды под колонией и вокруг нее.Фактически, он также образуется в мутантах, свободных от целлюлозы и курли, о чем можно судить по морфологическим признакам, таким как различная форма клеток, отсутствие или наличие деления клеток и образование жгутика в двух слоях (2). На самом деле, маленькие и яйцевидные клетки в верхнем слое растут медленно и, наконец, перестают расти, когда они перешли на физиологию стационарной фазы, основанную на RpoS, которая не только обеспечивает высокую стрессоустойчивость, но также является предпосылкой для образования локонов. и целлюлоза (39).Как только эти компоненты матрицы собраны, два слоя могут быть физически разрушены (см. Также фильмы S1 – S3 в дополнительном материале). Поскольку эта стратификация отражает две фундаментально разные физиологические стратегии бактерий – оптимизацию роста по сравнению с оптимизацией поддержания и выживания (39, 40) – и формирование градиентов питательных веществ связано с образованием любой биопленки, мы ожидаем, что двухслойная архитектура будет довольно общей. в бактериальных биопленках. Составляют ли голодные клетки верхний или нижний слой, на самом деле зависит от видоспецифичных метаболических свойств, в частности от того, необходим ли кислород для роста или нет (как обсуждается в ссылке 2).
Когда наши штаммы E. coli K-12 выращивают на бессолевой среде LB, экспрессия регулятора CsgD достаточно высока для образования высоких уровней локонов и целлюлозы, необходимых для образования гребней и морщин. Однако при выращивании бактерий на среде дрожжевой экстракт-казаминовые кислоты (YESCA) образование локонов и целлюлозы недостаточно для создания сложной морфологии (S. Herbst и R. Hengge, неопубликованные данные). Таким образом, такие условия удобны для изучения условий или мутаций, вызывающих появление морщин.В недавнем исследовании уропатогенного штамма E. coli UTI89, выращенного на среде YESCA, присутствие хлорида железа могло вызвать морщинистость макроколонии, которая позволяла плавать с верхнего слоя колонии. Это было интерпретировано как «бимодальное развитие популяции» на два отдельных слоя , индуцированное окислительным стрессом, хотя гены csgBAC и yaiC ( adrA ) не индуцировались в дальнейшем этим лечением (41). Было показано, что эти плавающие слои содержат клетки, которые были устойчивы к H 2 O 2 и экспрессировали curli (41), обе функции которых зависят от RpoS (39), и это открытие согласуется с физиологической двухслойной архитектурой, являющейся функцией градиент питательных веществ накапливается в макроколониях при всех условиях, а не вызывается окислительным стрессом de novo .Взятые вместе, это указывает на то, что окислительный стресс может специфически стимулировать образование целлюлозы и, следовательно, тканеподобную когезию и эластичность, необходимые для создания макроскопической структуры колонии, наблюдаемой DePas et al. (41), а также в нашем исследовании.
Целлюлоза – это четко локализованный архитектурный элемент, который образует «нанокомпозит» с завитыми волокнами и придает тканеподобные свойства.
На клеточном уровне волокна целлюлозы собираются в нерегулярные толстые нити и пластинчатые структуры, которые простираются между клетками (), а не плотно окружают клетки в виде «корзин», образованных амилоидными изогнутыми волокнами (2).Целлюлоза не распределена случайным образом в верхнем слое макроколонии (суммировано в). Скорее, массивное скопление вертикально расположенных целлюлозных волокон, листов и, возможно, даже длинных оболочек вдоль цепочек ячеек можно увидеть вблизи границы с нижним слоем, особенно в сильно изогнутой области кончиков гребней (до и) . Вдоль самой поверхности раздела формируются большие листы целлюлозы (). Кроме того, на поверхности колонии можно визуализировать композицию из целлюлозных и курчавых волокон (), на которой фактически невозможно различить два типа волокон.Этот матричный материал даже визуально сочетает в себе свойства изгибов и целлюлозы, поскольку он окружает клетки, но также обеспечивает нитевидные и листовые соединения между клетками, которые в целом создают относительно гладкую и сливающуюся защитную поверхность. Это может объяснить более раннее наблюдение, что матрица биопленки Salmonella , состоящая из локонов и целлюлозы, не переваривается ни целлюлазой, ни протеиназой К (12). Другими словами, E. coli и Salmonella естественным образом производят в своей матрице биопленки то, что ученые пытались оптимизировать, – целлюлозный «нанокомпозитный» материал (42, 43).Примечательно, что целлюлоза также может собираться вдоль жгутиков, что может быть обнаружено в переходной зоне между растущими и голодными клеточными слоями (), предполагая, что целлюлоза в целом легко собирается вдоль или вокруг нитчатых структур каркаса.
Локализация целлюлозы и образование гребней в макроколониях. (A) Схематическое резюме расположения целлюлозы и двухслойной архитектуры в макроколониях AR3110. Красный и оранжевый слои иллюстрируют зоны стационарной и постэкспоненциально-фазовой физиологии соответственно.Синими линиями обозначены зоны, богатые целлюлозой. (B) Схематическое изображение радиального расширения колонии. Круговые линии радиуса r, 2r и 3r представляют границу гипотетических колоний. Красные и синие линии в колониях с радиосигналом r и 2r, соответственно, имеют одинаковую длину, но разные углы кривизны, то есть бактерии, расположенные на этих линиях внешнего обода расширяющейся колонии, могут расти на последовательно меньшие области (показано заштрихованными красными штрихами). и синие области). Из-за «тканеподобной» сплоченности и эластичности колонии, которая генерирует как волокна завитков, так и целлюлозу, это приводит к изгибу колонии в радиально ориентированные гребни.
Только комбинация волокон целлюлозы и curli создает выраженную когезию, стабильность и эластичность макроколоний штамма AR3110, необходимую для развития сложной морфологии. Эластичность, в частности, может быть связана с большим количеством воды, содержащейся в целлюлозе (42). Напротив, матрица верхнего слоя с локонными волокнами только стабилизирует клетки на их месте, но является хрупкой и разрушает кольцевые структуры, типичные для деформации, вызывая только высокие уровни локонов (). С другой стороны, в макроколонии, которая производит только целлюлозу, сначала образуются небольшие морщинки, но затем скопление клеток, кажется, выталкивает все еще растущие клетки в изначально формирующиеся полости и со временем, кажется, разрастается морщинами ().В заключение, волокна curli действуют как клеточный «строительный раствор», который удерживает клетки на месте, тогда как целлюлоза придает эластичное сцепление этому клеточному «строительному материалу». Следует отметить, что недавнее сообщение приписало индуцированное этанолом или индуцированное диметилсульфоксидом повышенное сцепление и эластичность пленочных биопленок уропатогенных E. coli UTI89 с повышенной экспрессией curli (44, 45). К сожалению, в этих исследованиях целлюлоза не рассматривалась, и наши наблюдения за отдельными свойствами биопленки, обеспечиваемыми curli и целлюлозой, а также совместная регуляция обоих компонентов CsgD и c-di-GMP, предполагают, что биосинтез целлюлозы может быть стимулирован в параллельно с выражением curli в этих условиях.
Тканеподобные свойства целлюлозы позволяют создавать сложную трехмерную морфологию макроколоний
E. coli .Давно известно, что волокна curli и целлюлоза являются предпосылкой сложной морфологии макроколонии (12). Основываясь на наших исследованиях локализации с высоким разрешением, мы теперь можем ответить на вопрос, как комбинация целлюлозных и локонных волокон в определенных зонах матрицы E. coli создает эту конкретную трехмерную архитектуру.Чтобы решить этот вопрос, полезно концептуально деконструировать эту сложную морфологию – обобщенно называемую «рдар», «морщинистый» или «морщинистый» – на определенные элементы. К ним относятся размер и плоскостность макроколонии, а также отдельные элементы морфологической формы, такие как «гребни», «морщины» и «кольца», которые могут иметь радиальный, концентрический и / или неправильный характер.
Зрелые макроколонии AR3110 демонстрируют экстремальный размер и плоскостность ≥20 мм в диаметре, но только около 60 мкм в высоту на плоских участках, в отличие от макроколоний W3110 высотой> 200 мкм ().Вероятно, это функция целлюлозных волокон и листов, соединяющихся с curli-волокнами и жгутиками, которые, возможно, связывают клетки вместе, так что дочерние клетки внизу не могут просто накапливаться вертикально, а вынуждены распространяться латерально. Морщины и гребни возникают, когда макроколонии изгибаются, что возможно только из-за стабильности, когезии и эластичности композитной матрицы целлюлоза / завитки в верхнем слое макроколоний. Недавно было высказано предположение, что сморщивание макроколоний Bacillus subtilis в морщины происходит в областях локальной гибели клеток, что «обеспечивает выход для боковых сжимающих сил» (46).Хотя это исследование было сосредоточено на центральных и довольно старых регионах макроколоний, мы заметили, что образование гребней началось на внешних краях , а затем распространилось к более старому центру макроколоний E. coli (). Это указывает на объяснение, основанное на простой геометрии в сочетании с тканеподобными свойствами макроколонии. Когда растущая колония увеличивается в размере, участки одинаковой длины вдоль внешних краев колонии (показанные одинаковой длиной красной и синей линий в колониях с радиусом r и 2r дюйма) демонстрируют непрерывное уменьшение кривизны.Таким образом, хотя это кажется нелогичным, чем больше растет колония, тем меньше горизонтальное пространство, в которое могут расти клетки. В колонии с тканеподобной сплоченностью и эластичностью возникающее напряжение не может быть снято простым вертикальным складыванием одного потомства (обратите внимание, что формирование верхнего слоя с завитками и синтезом целлюлозы начинается довольно близко за внешним краем колонии [см. Рис. S3 в дополнительном материале]) (2). Скорее, весь регион колонии должен локально выпирать вверх.Получающееся в результате ослабление натяжения фактически позволяет областям между двумя вновь появившимися выступами снова сгладиться (). В каких местах действительно происходит коробление, кажется случайным и может зависеть от микрогетерогенности на поверхности агара.
Наши данные также показывают удивительные размеры хребтов. В зрелых макроколониях они могут быть> 4 мм в высоту, что соответствует> 30-кратной их ширине (около 120 мкм;). В «городе микробов» (47) эти гребни эквивалентны «небоскребам», демонстрируя тем самым превосходную стабильность и эластичность, которые придает нанокомпозитный материал целлюлоза / завитки.Подобные радиальные гребни обычно наблюдаются в биопленках макроколоний или пленок «недомашненных» бактерий. У грамположительных Bacillus subtilis их образование зависит от комбинации амилоид-образующего белка TasA и экзополисахарида (9, 10). Поскольку амилоиды распространены у бактерий (48, 49), нанокомпозиты амилоид-полисахарид в матрице могут стабилизировать сложную архитектуру биопленок у многих видов.
Физиологические роли целлюлозы и порождаемая целлюлозой морфология биопленок макроколоний.
По мере того, как мы начинаем понимать, как генерируется определенная архитектура макроколонии, возникает еще один интригующий вопрос: почему бактерии производят такую сложную морфологию. Какая польза от выращивания в очень больших, плоских и морщинистых колониях; зачем вкладывать все источники энергии и углерода в матрицу, позволяющую строить огромные хребты? Концентрация кислорода в макроколониях снижается до ок. 10% содержания кислорода в воздухе на глубине ∼60 мкм (41, 50). Поскольку морщинистость Pseudomonas aeruginosa или Bacillus subtilis вызывается нарушением дыхания, было высказано предположение, что увеличение поверхности за счет морщин облегчает доступ кислорода в качестве конечного акцептора дыхания (50, 51).Для E. coli мы предполагаем, что благоприятной адаптацией в отношении доступа к кислороду является плоскостность макроколоний, созданных матриксом curli / целлюлоза. Сгибание в морщины и гребни кажется вторичным и физическим следствием тканеподобных свойств этих очень плоских биопленок.
Кроме того, коробление определенных участков колонии имеет биологические последствия, поскольку оно относительно быстро перемещает большие группы клеток на большие расстояния, где они подвергаются воздействию новых условий микросреды.Например, во время образования гребня клетки, которые раньше находились в нижнем слое роста, быстро поднимаются до положения, в котором они подвергаются строгому голоданию. Эти клетки образуют тонкий слой внутри гребней, который не включает curli и выработку целлюлозы, что согласуется с тем, что гены CsgD и curli только слабо индуцируются, когда клетки подвергаются внезапному голоданию (например, в минимальной среде; E. Klauck and R Хенгге, неопубликованные данные). Также внизу хребта открывается новая зона роста.Из-за наибольшего расстояния от поверхности колонии можно ожидать, что эта зона будет иметь самое низкое содержание кислорода в макроколонии. Однако E. coli , которая может легко переключиться на анаэробный рост, полностью заполняет эти зоны ниже гребней (и до). Это контрастирует с ситуацией, недавно наблюдаемой для более зависимого от кислорода B. subtilis , где зоны под гребнями остаются открытыми и служат каналами, которые облегчают транспортировку жидкости (52).
С более экофизиологической точки зрения, рост в очень больших, плоских и похожих на ткани макроколониях во внешней среде – многие E.coli продуцируют curli и целлюлозу только при температуре ниже 30 ° C – это может обеспечить преимущество быстрого покрытия поверхностей, обеспечивающих питательными веществами, и, следовательно, лучшей конкуренции с бактериями, которые в большей степени зависят от кислорода для роста, такими как виды Bacillus или Pseudomonas . Кроме того, внешний слой колоний, покрытый нанокомпозитом curli / целлюлоза, может обеспечивать превосходную защиту от хищников или экзоферментов, продуцируемых конкурирующими видами, по сравнению с поверхностями, содержащими только завитки или целлюлозу.Кроме того, тканеподобное сцепление колоний может облегчить передачу из окружающей среды к хозяину и повысить выживаемость во время прохождения через желудок нового хозяина.
Целлюлоза также изменяет свойства волокон или нитей, с которыми она собирается, и дает дополнительные функции, не связанные с архитектурой колонии. Таким образом, он снижает опосредованную завитками адгезию к поверхностям (22). С другой стороны, он позволяет Salmonella прикрепляться к хитиновой поверхности гиф Aspergillus в смешанных биопленках (53) и контролирует прикрепление Rhizobium к корням растений (54).Целлюлоза также может противодействовать провоспалительному эффекту волокон curli при экспрессии в организме млекопитающего-хозяина (55, 56). Наконец, целлюлоза может ингибировать вращение жгутиков, что наблюдалось у мутанта, дефицитного по c-di-GMP-связывающему белку YcgR, белку, который останавливает вращение за счет прямого взаимодействия с базальным тельцем жгутика (57). Фактически, мы могли визуально продемонстрировать плотное переплетение целлюлозы со жгутиками в области перехода между слоями ростовой и неподвижной фаз макроколонии ().
Общее и дифференциальное регулирование амилоидных курлиновых волокон и целлюлозы и экологический контроль архитектуры макроколонии.
В настоящем исследовании мы сосредоточились на том, где в макроколонии встречаются целлюлоза и волокна локонов, и как эти компоненты матрицы вместе вносят вклад в формирование определенных морфологических элементов биопленок макроколонии. Фактически, морфогенез макроколоний регулируется в ответ на многие факторы окружающей среды, такие как присутствие соли (58), окислительный стресс (41), этанол (44) или влажность агаровой подложки.Например, макроколонии могут быть довольно жесткими только с несколькими очень высокими и радиальными гребнями, или они могут представлять собой целые «ландшафты» сложных морщин и большого количества высоких гребней (и). Очевидно, что не только местоположение как таковое , но также количество и, вероятно, соотношение между целлюлозными и локонными волокнами в определенных зонах макроколонии определяют жесткость и эластичность и, следовательно, имеют значение для создания определенной морфологии. На молекулярном уровне производство curli и целлюлозы совместно регулируется CsgD (21), что, в свою очередь, требует RpoS и нескольких ферментов, продуцирующих и разлагающих c-di-GMP, действующих в сложной регуляторной сети (26, 27, 59, 60). .На соотношение между двумя матричными компонентами может влиять дополнительный входной сигнал c-di-GMP при контроле производства целлюлозы (29).
Наконец, зоны с разными условиями в макроколонии не ограничиваются глобальной физиологической двухуровневой архитектурой. Фактически, образование макроструктур биопленки, таких как гребни или морщины, в свою очередь, порождает новые микросреды. В будущем будет интересно посмотреть, насколько пространственно модулирует передача сигналов c-di-GMP множественными DGC и PDE синтез curli, целлюлозы и, возможно, дополнительных матричных компонентов и тем самым контролирует архитектуру биопленки в ответ на макро- и условия микросреды.
Архитектурные элементы: как создать действительно красивую лестницу
Просмотрите доску вакансий Architizer и подайте заявку на должности архитектора и дизайнера в некоторых из лучших мировых фирм. Нажмите здесь, чтобы подписаться на нашу рассылку новостей о вакансиях.
В некоторых проектах лестница – это просто аллея, обеспечивающая вход или выход для жителей. В других случаях это скульптурные, резкие, минималистичные или завораживающие архитектурные элементы, которые могут придать проекту неповторимый характер.Если вы архитектор, который верит в то, что лестница должна быть максимальной, рассмотрите этих специализированных производителей и их инновационные продукты для лестниц – от отдельных компонентов до полных систем – для своего следующего проекта.
Glass Treads от Nathan Allan Glass Studios могут создавать неземной вид, а также пропускать свет в пространство внизу.
Nathan Allan Glass Studios: Стеклянные ступени
Специалист по стеклу предлагает ступени и пол с двумя различными вариантами поверхности: наждачная бумага для стекла создает дополнительное сцепление, наплавляя частицы стекла на поверхность ходьбы, а покрытие Silken обеспечивает сцепление за счет рисунков с слегка протравленным покрытием.Оба они прошли лабораторные испытания на соответствие требованиям ADA по коэффициенту скольжения.
EeStairs предлагает эту балюстраду с разным уровнем открытости для удовлетворения различных требований по дизайну и безопасности.
EeStairs®: Органическая балюстрадная система Cells
Cells от EeStairs – это замысловатая балюстрада с органическими очертаниями. Система может быть адаптирована для прямых или винтовых лестниц, с меньшими или большими ячейками и в любом цвете RAL.
Материал Cosentino Dekton очень прочен для пешеходов и хорошо очищается.
Косентино: Dekton
Ультракомпактная поверхность состоит из стекла, фарфора и кварца, которые прошли запатентованную технологию воздействия высоких температур и давления (так называемая технология спекания частиц). Этот прочный, простой в уходе и чистоте материал, подходящий для различных целей, от внешней облицовки до столешниц, является отличной альтернативой ступеням из камня, металла, дерева или бетона, поскольку он может имитировать все вышеперечисленное.
CRL-U.Система перил SRS от S. Aluminium создает шикарную внутреннюю или внешнюю лестничную балюстраду.
CRL-U.S. Алюминий: система перил SRS
Хорошо подходит как для коммерческих, так и для жилых проектов, эта система стеклянных перил может быть установлена на большинстве одобренных оснований и произведена в соответствии с требованиями строительных норм или даже превосходящими их. Фурнитура Standoff может быть завершена плоскими / плоскими или куполообразными крышками, чтобы дополнять различные стили проекта.
Лестница Marretti’s Cloud сочетает кориан, нержавеющую сталь и стекло.
Марретти: Облако
Последняя модельMarretti отличается проступью Corian со вставкой из нержавеющей стали и перилами из сверхпрозрачного структурного стекла. Corian доступен в дополнительных цветах, а также может быть заменен на дерево или смолу с другой отделкой и цветом.
Плавающая лестница от EeStairs
EeStairs: Плавающая лестница
Система «Плавающая лестница» прикрепляет ступеньки к стене с одной стороны. Хотя другую сторону можно оставить без опоры, существует множество вариантов добавления рельса.Ступени могут быть выполнены из дерева, стекла, камня или других материалов.
Кронштейны для перил Halliday + Baillie
Halliday + Baillie: Кронштейны для перил
Предлагаемые из алюминия или нержавеющей стали кронштейны для лестничных перил Halliday + Baillie могут быть прикреплены к стенам, стеклу или панелям. Блоки бывают круглого и квадратного стилей, различных размеров, а также скрытого или открытого оборудования. Модель HB 525 также может содержать проводку для подрельсового освещения.
Балтийская сетка GKD, используемая для ограждения и экранирования винтовой лестницы
GKD-USA: Балтика
Металлические тканиGKD часто используются для фасадов, экранов, защиты от солнечных лучей, крышек колонн, потолков и стен. Но прочный материал также идеально подходит для лестничных клеток. Показана сетка Baltic, которая состоит из нержавеющей стали AISI Type 316 и имеет открытую площадь 38 процентов.
Bisca создает свои индивидуальные лестницы из различных материалов, таких как зеленый дуб (слева), орех (справа) и стекло.
Bisca: Bespoke Staircases
Британская компания Bisca предлагает индивидуальные лестницы любого стиля, от прямых до консольных и винтовых, от традиционных до деревенских или современных. Также можно полностью настроить материалы – от дерева до стекла – при условии, что они соответствуют нормам.
Просмотрите доску вакансий Architizer и подайте заявку на должности архитектора и дизайнера в некоторых из лучших мировых фирм. Нажмите здесь, чтобы подписаться на нашу рассылку новостей о вакансиях.
Мы не можем найти эту страницу
(* {{l10n_strings.REQUIRED_FIELD}})
{{l10n_strings.CREATE_NEW_COLLECTION}} *
{{l10n_strings.ADD_COLLECTION_DESCRIPTION}}
{{l10n_strings.COLLECTION_DESCRIPTION}} {{addToCollection.description.length}} / 500 {{l10n_strings.TAGS}} {{$ item}} {{l10n_strings.PRODUCTS}} {{l10n_strings.DRAG_TEXT}}{{l10n_strings.DRAG_TEXT_HELP}}
{{l10n_strings.ЯЗЫК}} {{$ select.selected.display}}{{article.content_lang.display}}
{{l10n_strings.AUTHOR}}{{l10n_strings.AUTHOR_TOOLTIP_TEXT}}
{{$ select.selected.display}} {{l10n_strings.CREATE_AND_ADD_TO_COLLECTION_MODAL_BUTTON}} {{l10n_strings.CREATE_A_COLLECTION_ERROR}}Использование архитектурных элементов для контраста
Художник из Сан-Диего Дэнни Гриего считает, что включение «городской линии» в его картины делает их намного сильнее – и имитирует то, как многие из нас видят мир.Как так?
Изображение в свинце: «9’-2», Дэнни Гриего, масло, 9 x 12 дюймов
«Мне очень нравится рисовать городские элементы», – говорит Гриего. «Мне нравится, как они помогают формировать и определять картину, особенно когда в нее примешивается природа. Меня особенно интересует то, как искусственные конструкции интегрируются с природой ».
Художник очень четко понимает, какое влияние на его искусство играют городские линии. «Использование городских линий привлекает внимание к моим картинам», – говорит Гриего. «В моих картинах преобладает городская линия, то есть они более резкие и четкие, чем органические линии природы.Они необходимы, как прямая линия на рисунке или портрете, помогает поддерживать и структурировать. Эти два аспекта, использование городских линий и органических линий природы, могут создавать в картине интересные формы или абстракции ».
«Городская дымка», Дэнни Гриего, масло, 20 x 16 дюймов,Он объясняет: «Например, большое открытое пространство, такое как небо, может занимать большую часть картины, но может контрастировать с архитектурным элементом. Архитектурные элементы могут быть большими или маленькими, и внутри этих форм может происходить множество тонких и интересных вещей – другие формы, изменения значений, изменения цвета и т. Д.Это верно в двух моих картинах «9’-2» и «Городская дымка». Архитектурные элементы являются доминирующими, в значительной степени силуэты по стоимости, но городская линия, которую они создают на фоне неба, мне интересна: очень прямая, темная и высококонтрастная. Сделано руками человека. Использование городской линии в этом контексте – потому что она так сильно контрастирует с небом – создает тот знакомый вид или ощущение, которое возникает, когда проезжаете под мостом на автостраде или видите рекламные щиты на расстоянии. Это кусочек городского “.
«Pacific Highway Glow» Дэнни Гриего, масло, 12 x 24 дюйма.Griego чувствует себя комфортно, когда любой элемент – природа или городская структура – занимает первенство в композиции. «« Свечение Тихоокеанского шоссе »и« Марстон Хаус »демонстрируют использование городских линий, но не небо, занимающее большую часть картины, а архитектура, которая доминирует», – отмечает он. «Негативные формы неба вторичны, но все же есть игра с абстракцией. Архитектура – звезда шоу, а мягкие органические формы природы хорошо ее дополняют. В этих двух картинах мне нравится концепция «возвышения».«Для меня это означает, что точка обзора находится снизу и смотрит немного вверх. Использование городских линий в этом режиме с немного преувеличенной перспективой может заставить зрителя почувствовать, что он втягивается в картину. И если он хорошо читается через всю комнату, значит, вы знаете, что добились некоторого успеха ».
«След с Керлью-стрит» Дэнни Гриего, масло, 9 x 12 дюймов.До сих пор Григо использовал городские элементы в качестве сильных игроков в картинах, независимо от того, являются ли они первичными или второстепенными по отношению к композиции.Но он также использует предметы меньшего размера, чтобы придать пейзажу немного привлекательности. «Городские линии на картине также могут быть тонкими и могут быть помещены в основную ценность или массу картины», – говорит Григо. «Мне нравится, например, как телефонные столбы создают точки, в которых они крепятся к земле. Это элемент дизайна, который для меня создает арматуру на картине, особенно когда есть еще одна линия, прикрепленная по диагонали к столбу от земли, которая образует треугольник.
«Дом Марстона» Дэнни Гриего, масло, 12 x 16 дюймов.«В« Urban Trail Head »вы можете увидеть пример этого. В «Trail off Curlew Street» искусственный элемент шеста и линии от земли до шеста расположены под противоположными углами к наклонному ландшафту. Здесь создается еще один треугольник. Без этого прямого столба под определенным углом картина не могла бы держаться вместе. Дом слева – еще одна возможность противопоставить прямые линии природе, что добавляет разнообразия и разрушает в основном органическое ощущение этой картины.”
«Городская тропа» Дэнни Гриего, масло, 9 x 12 дюймов.Очевидно, эта предрасположенность к включению городских элементов в пейзажи заложена в художественном мозгу Григо. «Я не совсем понимаю, почему, но я постоянно ищу это, когда выхожу из дома», – говорит он. «Я еду и делаю мысленные пометки для будущих картин, основанных на этом. Надеюсь, это безопаснее, чем писать текстовые сообщения и водить машину! ”
Идентификация архитектурных элементов в существующих системах
Саймон недавно говорил о разрыве между архитектурой программного обеспечения и кодом и о том, как закрыть это с помощью архитектурно очевидного кодирования.Он также создает инструменты, позволяющие выразить архитектуру программного обеспечения в виде кода.
Если вы работаете над новым проектом, то включение аннотаций для помощи в навигации – отличное решение, но что, если вы унаследовали большую систему с пробелом в коде модели? Или если вы только на каком-то этапе проекта понимаете, что вам не хватает модели, которая помогла бы вам понять его растущую сложность? Ну, Саймон также подумал о сканировании аннотаций Spring для предоставления этих данных. Это работает довольно хорошо, и это заставило меня задуматься о других артефактах в коде, которые можно извлечь для этих диаграмм.
(В более формальных терминах – для тех из вас, кто любит цитировать ISO42010 – мы пытаемся извлечь архитектурный элемент из кода, который может быть отображен в пределах архитектурных представлений . Конечно, элементы могут быть из множества различных абстракций, уровней и детализации и поэтому должны быть помещены в разные точки зрения.)
Итак, что мы можем извлечь из существующей / устаревшей системы, чтобы получить представление о ней? Некоторые предложения включают: